
| [ начало ] | [ Б ] |
Бесхвостые гады
или Батрахии (Batrachia, см. табл. Бесхвостые гады I, II и III) — первый и высший отряд класса Земноводных, или Голых гадов (Amphibia).
БЕСХВОСТЫЕ ГАДЫ (BATRACHIA). I.
1. Пипа суринамская. 2. Зеленая или снедная лягушка. 3. Бурая лягушка. 4. Превращения лягушки. 5. Повитушка или жаба-акушерка. 6. Рогатая лягушка. 7. Жерлянка. 8. Жаба обыкновенная. 9. Летучая квакша. 10. Обыкновенная квакша или древесница. 11. Сумчатая лягушка.
Это самый богатый видами порядок земноводных: из общего числа всех представителей класса (169 родов и 917 видов) к нему принадлежат около 138 родов с 800 видами; из этого числа в Европейской России водятся 5 родов с 8 видами и один вид в Сибири, а в Европе вообще 8 родов с 12 видами. Главнейшие отличительные признаки Б. гадов (см. табл. Бесхвостые гады I) следующие: тело во взрослом состоянии всегда бесхвостое, с четырьмя более или менее развитыми ногами, из коих задние всегда длиннее передних; жабр во взрослом состоянии нет; дышат легкими и кожей. Короткое, более или менее плоское тело этих животных разделяется на голову, туловище и конечности. Голова плоская, широкая, прямо переходящая в туловище, так что шеи у Б. гадов снаружи не заметно. На голове, именно по бокам ее передней части, помещаются большие глаза, которые могут выпячиваться и втягиваться; раек их всегда имеет яркую, большей частью золотистую окраску; глаза защищены веками и мигательной перепонкой; последняя прозрачна и может закрывать весь глаз; только у пипы нет век. Маленькие ноздри (Б. Г., табл. II, рис. 11) находятся спереди, на конце морды, открываются почти вертикально книзу в ротовую полость и могут закрываться особыми клапанами (складками кожи).
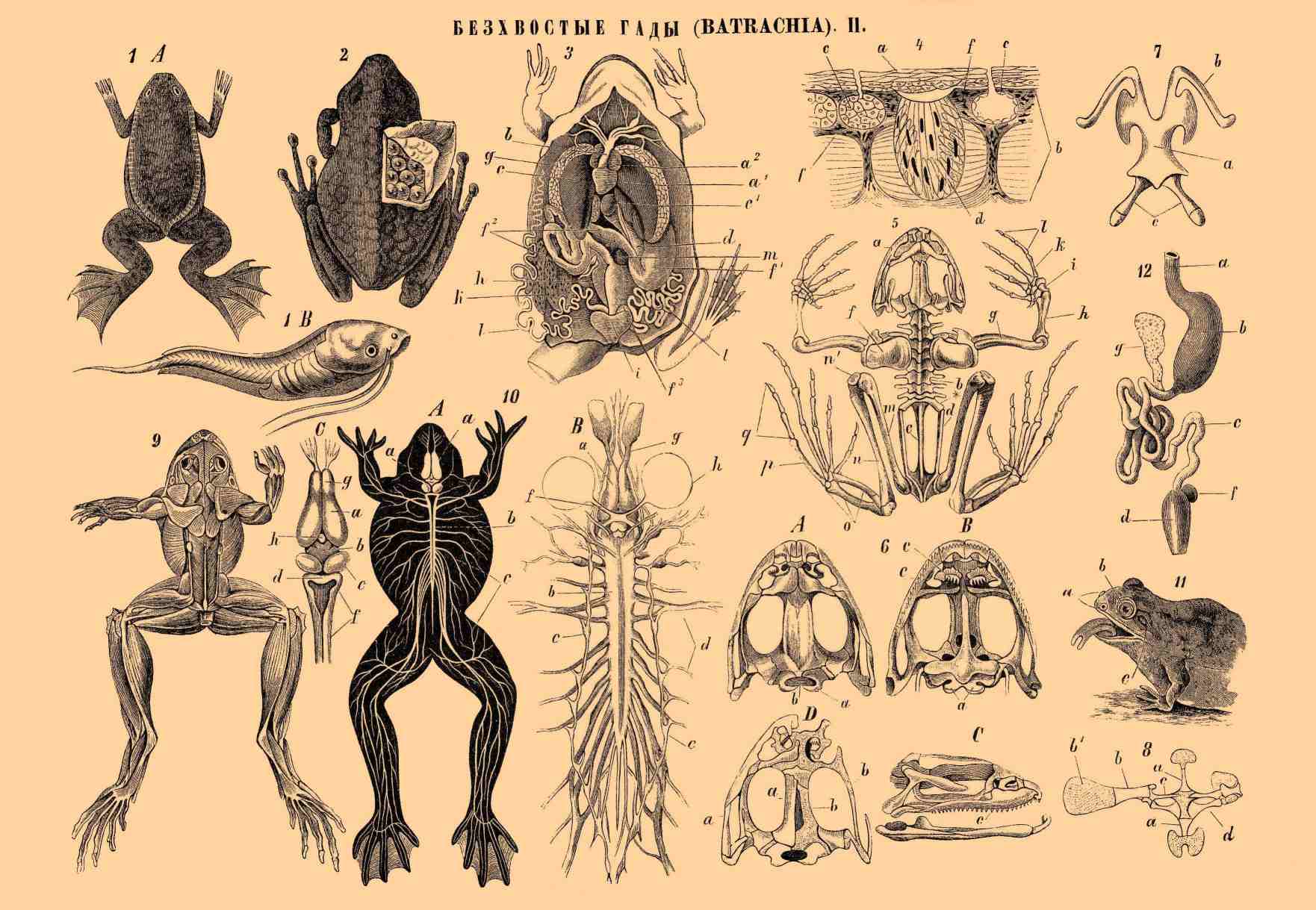
БЕСХВОСТЫЕ ГАДЫ (BATRACHIA). II.
Рис. 1. Дактилера капская (Dactylera capensis). A. взрослое состояние. B. Личинка (головастик). Рис. 2. Спинномешочница (Notodelphys ovifera); с одной стороны кожа спины разрезана и отворочена в сторону, чтобы можно было видеть спинную суму с заключающимися в ней яичками. Рис. 3. Общее расположение органов в теле зеленой лягушки (Rana esculenta): a — сердце: a1 — желудочек сердца, a2 — предсердие, b — кровеносные сосуды (артериальные стволы), сообщающиеся с сердцем; c — печень; c1 — желчный пузырь; d — поджелудочная железа; f — пищеварительный аппарат: f1 — желудок, f2 — тонкая кишка, f3 — толстая кишка; g — легкое; h — почка; i — мочевой пузырь; k — яичник; l — яйцепровод, m — селезенка. Рис. 4. Поперечный разрез кожи лягушки: a — кожица (epidermis), b — собственно кожа (cutis sive derma), c — маленькая кожная железка; d — большая кожная железка; f — хроматофор (пигментная клетка). Рис. 5. Скелет зеленой лягушки (Rana esculenta), сзади: a — череп, b — позвоночный столб, c — крестец, d — тазовая кость, f — лопатка, g—l — кости передней конечности: g — плечевая кость, h — кости среднеплечия, или предплечья, i — кости запястья, k — кости пясти, l — суставы пальцев; m—q — кости задней конечности: m — бедренная кость, n — кости голени, o — кости пятки, p — плюсневые кости, q — суставы пальцев. Рис. 6. Череп зеленой лягушки (Rana esculenta). A — сверху, B — снизу, C — сбоку: a — мыщелок затылочной кости, b — затылочная дыра, c — зубы; D — череп сверху, слева изображены отдельные кости (a), а справа кости черепа сняты и видны лежащие под ними хрящи (b), которые составляют первичный хрящевой череп. Рис. 7. Подъязычная кость обыкновенной жабы (Rufo vulgaris): a — тело подъязычной кости, b — рога подъязычной кости, c — остатки жаберных дуг. Рис. 8. Грудина и плечевой поясок зеленой лягушки (Rana esculenta): a — грудина или грудная кость, b — лопатка, b1 — надлопаточный хрящ, c — ключица, d — воронья кость. Рис. 9. Мышцы (мускулатура) зеленой ягушки (Rana esculenta). Рис. 10. Нервная система зеленой лягушки (Rana esculenta): A — головохребетная нервная система в теле животного, сверху a — головной мозг, b — спинной мозг, c — нервы. B — Головохребетная и симпатическая нервная система, выделенная из тела, снизу: a — головной мозг, b — спинной мозг, c — узлы симпатической нервной системы, d — нервы спинного мозга, f — зрительный нерв, g — обонятельный нерв, h — глаз. C — отдельный головной мозг: a — полушария головного мозга, b — промежуточный мозг, или зрительные бугры, c — средний мозг, или двухолмие, d — малый мозг, или мозжечок, f — продолговатый мозг, g — обонятельная долька, h — так называемая мозговая железка (третий зрительный центр). Рис. 11. Голова бурой лягушки (Rana temporaria) сбоку, с открытым ртом: a — ноздря, b — глаз, c — барабанная перепонка (слуховой орган). Рис. 12. Пищеварительные органы бурой лягушки (Rana temporaria): a — пищепровод, b — желудок, c — тонкая кишка, d — толстая кишка, f — селезенка, g — поджелудочная железа.
По бокам задней части головы помещается большая круглая барабанная перепонка; она покрыта кожей и у одних Б. гадов ясно просвечивает через нее, а у других почти незаметна. За весьма немногими исключениями (жабы, Bufo; Пипа, Pipa) все Б. гады имеют зубы, расположенные на верхней челюсти и на небе; нижняя челюсть обыкновенно беззубая; небные зубы располагаются двумя короткими поперечными рядами между носовыми отверстиями или позади них. Зубы мелки и у всех европейских видов зазубренные или многодольчатые, причем зазубрины и дольки их всегда загнуты внутрь. Рот широкий, и ротовая щель оканчивается позади глаз. Язык довольно большой, по большей части толстый и мясистый, часто спереди приросший, а сзади свободный, так что может выбрасываться изо рта и служит органом для схватывания добычи; реже он срощен с дном ротовой полости (напр. у пипы). Туловище более или менее плоское. Конечности у всех хорошо развиты; задняя пара ног сильнее и длиннее передней, которая всегда согнута внутрь. На передних ногах явственно развиты 4 пальца, которые обыкновенно свободны и имеют приблизительно одинаковую длину; пятый, большой палец, скрыт под кожей в виде маленького хрящика или маленькой косточки, так что снаружи большим пальцем кажется обыкновенно второй. На задних ногах по большей части 5 пальцев, которые бывают неодинаковой величины (обыкновенно четвертый палец самый длинный, большой — самый короткий) и по большей части соединены между собой половинными или полными плавательными перепонками; у многих на внутреннем крае пятки находится бугорок, называемый пяточным бугорком, или шестым пальцем. Лишь у немногих Б. гадов на пальцах бывают когти (только у Dactylethra, таблица II, рис. 1), но у многих на нижней поверхности их находятся особые подушечки, или бородавка, или шишкообразные утолщения. Кожа Б. гадов всегда голая и б. ч. не имеет никаких твердых эпидермоидальных образований; напротив, вследствие сильного развития желез (см. таблица II, рис. 4) она бывает скользкой и гладкой (напр., у лягушки); часто же неровной и покрытой бородавками, именно в тех случаях, когда выделяются острые и едкие жидкости (напр., у жаб, табл. I, рис. 8). В последнем случае особые железы скучиваются в большом количестве на определенных местах, преимущественно около уха, и образуют сильно выдающиеся железистые возвышения; подобные железистые скопления встречаются также на голенях (напр., у жабы камышовой, Bufo calamita) и по бокам тела. Кожа Б. гадов (табл. II, рис. 4) повсюду богата нервами и сосудами, тонка, и потому она не только весьма чувствительна, но играет важную роль при дыхании (кожное дыхание — perspiratio).
Из внутренних органов наибольшие особенности представляет скелет (табл. II, рис. 5), состоящий из черепа, позвоночного столба и костей конечностей и поясков, прикрепляющих конечности к позвоночному столбу. Череп (табл. II, рис. 6) состоит из очень маленькой черепной коробки и из очень развитого, дугообразно расположенного челюстно-небного аппарата. Квадратная кость, служащая для сочленения нижней челюсти с черепом, неподвижно соединена с черепом, небные, межчелюстные и челюстные кости также неподвижно срастаются с черепной коробкой, так что подвижна только одна нижняя челюсть, обе половины которой прочно соединены между собой. Череп соединяется с первым шейным позвонком посредством двух мыщелков. Череп Б. Г., как и у всех земноводных вообще, костяной и состоит из многих отдельных костей, но под ним сохраняется первичный, или зародышевый, хрящевый череп, который состоит из многих отдельных хрящей. Подъязычная кость (табл. II, рис. 7) большая, снабжена двумя рогами и многими отростками. Позвоночный столб состоит из 10, а у некоторых вследствие срастания отдельных позвонков из 9 и 8 отдельных позвонков, именно из 9 собственно туловищных и одного очень длинного хвостового позвонка. Последний туловищный позвонок, или крестцовая кость, соединяется посредством больших поперечных отростков (крестцовые отростки) с седалищными костями; тазохвостовой позвонок, или копчиковая кость, не есть одна кость, а представляет сращение многих отдельных хвостовых позвонков. Тела позвонков спереди вогнутые, а сзади выпуклые; форма крестцовых поперечных отростков, которые бывают то узкими и цилиндрическими (напр., у лягушек, Rana), то плоскими и расширенными (напр., у повитушки, Alyt e s, пипы, Pipa), имеет диференциальное значение для систематики Б. гадов. Ребер совсем нет, но зато поперечные отростки туловищных позвонков достигают значительной длины. Грудина (табл. II, рис. 8) хорошо развита. Плечевой и тазовый пояски имеются у всех; первый (табл. II, рис. 8) характеризуется значительной величиной плоских лопаток и крепким соединением с грудной костью; кроме лопаток, в состав этого пояска входят еще две ключицы и две вороньи кости; тазовый поясок (табл. II, рис. 5) образуется тремя парами костей — седалищными, подвздошными и лобковыми — и характеризуется длинными и узкими подвздошными костями; у некоторых Б. Г., напр., у лягушек, подвздошные и лобковые кости срастаются между собой в две подвздошно-лобковые кости. На передних конечностях лучевая и локтевая кость сращены друг с другом в одну предплечную кость, точно так же как и большая и малая берцовые кости задних конечностей сращены в одну кость голени; пяточные кости удлинены. Мускулатура (табл. II, рис. 9) очень сильно развита, и есть много подкожных мускулов. Нервная система (табл II, рис. 10) состоит из головного и спинного мозга и из симпатической нервной системы. Головной мозг состоит из двух полушарий большого мозга, промежуточного и среднего мозга, мозжечка и из продолговатого мозга. Полушария большого мозга довольно развиты, и спереди их находятся обонятельные доли; промежуточный и средний мозг обособлены, и последний состоит из двух возвышений (двухолмие); мозжечок имеет вид очень маленькой и тоненькой пластинки; продолговатый мозг сильно развит. Число черепных нервов меньше, чем у высших животных. Спинной мозг по объему значительно превышает головной и выполняет позвоночный канал на всем его протяжении. Спинной мозг дает б. ч. 10 пар нервов. Осязание развито хорошо, и главным органом этого чувства служит тонкая, богатая нервами кожа. Всегда ли развито чувство вкуса — неизвестно; хотя на языке и были найдены вкусовые сосочки, но Б. Г. проглатывают пищу неизмельченной, а язык (табл. II, рис. 11) часто служит у них для других отправлений, напр., для ловли добычи, а у некоторых (Безъязычные) обособленного языка нет. Органы обоняния состоят из двух снабженных складками слизистой оболочки носовых полостей, сообщающихся с ротовой полостью и открывающихся кнаружи ноздрями (табл. II, рис. 1 1а). Слуховой аппарат состоит из внутреннего уха, т. е. твердого лабиринта, заключающего в себе слуховой мешок, и из трех полукружных каналов и из среднего уха, или барабанной полости, которая ограничивается кнаружи барабанной перепонкой (табл. II, рис. 1 1 с) и сообщается с полостью глотки посредством Евстахиевой трубы. В среднем ухе помещается только одна слуховая косточка, пирамидка; улитки нет. У всех Б. Г. есть глаза (табл. II, рис. 11в), представляющие ту особенность, что имеют особую мышцу, при помощи которой глазное яблоко может глубоко втягиваться. Пищеварительный аппарат (табл. II, рис. 12) представляет малое развитие, и вообще пищеварительный канал немного превышает длину тела. Ротовая полость широкая, у большей части Б. гадов вооруженная зубами, сидящими в верхнечелюстных, межчелюстных и небных костях и служащими лишь для схватывания пищи, но не для размельчения ее. У многих ротовая полость сообщается с горловыми мешками, которые бывают одиночными или парными и открываются в ротовую полость отверстиями, расположенными позади языка; через эти отверстия мешки наполняются воздухом и служат для усиления звука, почему и называются голосовыми мешками. Собственно пищеварительные органы состоят из пищепровода, желудка, тонких и толстых кишок (табл. II, рис. 12). Пищепровод короткий и широкий; желудок хотя слабо, но заметно отличается от остального пищеварительного канала; тонкая кишка представляет мало извилин; более широкая толстая кишка ведет в клоаку, имеющую продолговато-круглое наружное отверстие. Слюнных желез нет; печень, желчный пузырь и поджелудочная железа всегда есть, и первая разделена на 2 дольки (табл. II, рис. 3 и 12). Все гады во взрослом состоянии дышат легкими (табл. II, рис. 15). Легких — два; они одинаково развиты и суть два мешка с большой полостью и с сетчатыми стенками. Дыхательное горло очень коротко, так что оба бронха (ветви дыхательного горла) почти непосредственно примыкают к гортани, состоящей из нескольких хрящей и имеющей хорошо развитые голосовые связки. Вдыхание и выдыхание воздуха при закрытом рте происходит через носовые отверстия; при вдыхании действует мускулатура подъязычной кости, а при выдыхании — мускулатура брюшной стенки. Кроме легких, в дыхании принимает деятельное участие и кожа. Сердце (табл. III, рис. 13) состоит из одного желудочка и двух предсердий. В желудочке сердца артериальная и венозная кровь смешиваются, хотя и не вполне. Селезенка (табл. II, рис. 12f) маленькая, шарообразная; она помещается вблизи толстой кишки. Лимфатическая система (табл. III, рис. 14) состоит из лимфатических сосудов и, кроме того, имеет ритмически пульсирующие лимфатические сердца, которых большей частью бывает две пары: одна в плечевой области, другая по бокам крестца. Лимфатических железок нет. Почки (табл. III, рис. 16d и f) лежат по бокам задней части позвоночного столба. Выходящие из почек каналы (мочеполовые протоки) открываются в клоаку; есть также и мочевой пузырь, но он не сообщается с мочеточниками, а открывается в клоаку и из нее наполняется мочой. Б. гады, как и все земноводные вообще, раздельнополы, только у некоторых из них (напр. у жаб, у чесночниц) у самцов находятся следы гермафродитизма (табл. II, рис. 18). Женские половые органы (табл. III, рис. 17) состоят из двух более или менее извитых яйцепроводов, оканчивающихся в клоаку. Каждый яйцепровод имеет два отверстия — переднее, широкое, воронкообразное, обращенное в полость тела, и заднее, обращенное в клоаку.

БЕСХВОСТЫЕ ГАДЫ (BATRACHIA). III.
Рис. 13. Кровеносная система лягушки бурой (Rana temporaria): a — легкое, b — сердце, c — артериальный ствол, d — легочная артерия, f — аортная дуга, g — ствол аорты, h — сонная артерия, i — легочная вена, k — нижняя полая вена. Рис. 14. Лимфатическая система зеленой лягушки (Rana esculenta): A — Лимфатическая система кожи: a — лимфатические сосуды. B — лимфатические сердца: BI — передняя часть тела лягушки с передними лимфатическими сердцами (а), BII — задняя часть тела лягушки с задними лимфатическими сердцами. Рис. 15. Дыхательный аппарат зеленой лягушки. A — гортань с легочными мешками: a — гортань; B — весь дыхательный аппарат: a — подъязычная кость, b — гортань, c — бронх, или ветвь дыхательного горла, d — легкое. Рис. 16. Мужской мочеполовой аппарат зеленой лягушки (Rana esculenta): a — жировое тело, b — семенник, или мужская половая железа (testiculus), c — семенные канальцы, d — верхняя часть почки, в которую оканчиваются семенные канальцы (Вольфово тело), f — остальная часть почки (настоящая почка), g — мочеполовой канал, h — семенной пузырек, i — клоака (вскрытая), k — нижний конец прямой кишки, отогнутый в сторону, l — мочевой пузырь, отогнутый в сторону. Рис. 17. Женский половой аппарат зеленой лягушки: a — яичник (ovarium), b — яйцепровод, b1 — нижний, расширенный конец яйцепровода, c — клоака. Рис. 18. Мужской мочеполовой аппарат жабы (Bufo vulgaris): a — жировое тело, b — почка, c — мочеточник, d — семенник, f — зачаточный яичник, g — семяпровод, h — мочеполовой канал. Рис. 19. Икра зеленой лягушки (Rana esculenta), прикрепленная к листьям водяного растения. Рис. 20. Дробление желтка и развитие зародыша в яйце зеленой лягушки: a — яйцо, b—f — различные стадии дробления желтка, g—i — зародыш. Рис. 21. Превращения чесночницы (Pelobates fuscus): a — часть яйцевого шнурка, a1 — отдельное увеличенное яйцо с зародышем, b — несколько молодых головастиков, сидящих на ветке водяного растения, в естественную величину, c — безногий головастик с наружными жабрами, снизу, d — головастик более поздней стадии развития, имеющий хвост и пару задних ног, g — головастик с хвостом и двумя парами ног, h — молодая чесночница (Pelobates fuscus) еще с хвостиком, i — вполне развитая чесночница (без хвостика). Рис. 22. Зародыш и головастик зеленой лягушки (увеличен). A — зародыш: a — жаберные щели; B — головастик с наружными жабрами, сбоку: a — жабра, c — присоска; C — то же, снизу: b — роговые челюсти, c — присоска; D — головастик более поздней стадии развития, со внутренними жабрами, сбоку: b — роговые челюсти, c — присоска, d — жаберная крышка, из-под которой видна часть жабр. Рис. 23. Череп головастика зеленой лягушки (хрящевой, или первичный, череп). Рис. 24. Превращения подъязычной кости. A — подъязычная кость зеленой лягушки, B и C — подъязычная кость головастиков лягушки в двух стадиях развития: a — тело подъязычной кости, b — рога подъязычной кости, c — жаберные дуги, d — задние отростки подъязычной кости. Рис. 25. Мускулы головастика зеленой лягушки (Rana esculenta). Рис. 26. Пищеварительные органы головастика зеленой лягушки. A — головастик снизу, со вскрытою полостью тела, в которой помещается пищеварительный канал, спирально свернутый в виде часовой пружины. B — пищеварительный аппарат головастика лягушки, отчасти высвобожденный из тела и развернутый: a — пищепровод, b — желудок, c — кишечный канал. Рис. 27. Органы кровообращения головастика зеленой лягушки с наружными жабрами: a — жабра, b — главный артериальный ствол, c — жаберная артерия, d — жаберная вена, f — общая жаберная вена, или выносящий сосуд (arteria epibranchialis), g — ствол аорты. Рис. 28. Органы кровообращения головастика зеленой лягушки со внутренними жабрами и с легкими: a — жабра, b — артериальный ствол, c — жаберная артерия, d — общая жаберная вена, f — ствол аорты, g — сердце, h — легкое, i — легочная артерия, k — легочная вена, l — нижняя полая вена. Рис. 29. Развитие легких у головастика зеленой лягушки: a — легкие, b — бронхи, или ветви дыхательного горла, c — глотка, вскрытая, d — пищепровод, f — подъязычная кость. На этом рисунке видно, что легкие и бронхи образуются как выпячивания по бокам нижнего конца глотки.
Стенки яйцепроводов выделяют студенистое вещество, облекающее яички, которые выходят наружу в виде икры. К заднему концу каждого яйцепровода присоединяется выходящий из почки мочеточник. Мужские половые органы (табл. III, рис. 16) состоят из двух семенников, из выводных семенных канальцев, двух мочеполовых каналов и из двух семенных пузырьков. Семенники — продолговато-круглые тела, помещающиеся вблизи почек и покрытые дольчатым жировым телом. Из них выходят многие выводные семенные канальцы, внедряющиеся в передний отдел почки. Из каждой почки выходит мочеполовой канал, открывающийся в клоаку. Сбоку нижнего конца каждого мочеполового канала находится по продолговатому мешочку — семенному пузырьку, в котором временно сохраняется мужское семя, поступающее в него из мочеполового канала, выносящего также и мочу. Мужское семя выпрыскивается из клоаки на икру в момент ее выхождения из тела самки. У жаб и чесночниц самцы имеют зачаточные яичники, помещающиеся вблизи семенников. Эти яичники не функционируют (табл. III, рис. 18). — Самцы отличаются от самок большей частью более громким голосом, который у многих усиливается еще голосовыми мешками; присутствием на большом пальце особого утолщения, так называемой пальцевой мозоли; иногда более развитыми плавательными перепонками, а у некоторых половые различия выражаются также окраской и величиной, причем самцы бывают обыкновенно ярче окрашены и меньше самок.
Размножение — половое. Спаривание происходит главным образом весной. При этом самец с силой обхватывает самку передними ногами за туловище и поливает своим семенем яйца, выходящие вследствие давления из самки в виде шнуров (у жаб) (табл. III, рис. 21а) или в виде комков (у лягушек, табл. III, рис. 19). Некоторые Б. гады проявляют известную заботливость о своем будущем потомстве. Так, у повитушки (Alytes obstetricans, табл. I, рис. 5) самец наматывает большие, гроздевидно соединенные между собой яйца на свои задние ноги, закапывается и уже позже относит яйца в воду, перед самым вылуплением из них детенышей. У пипы (Pipa surinamensis, табл. I, рис. 1) самец по оплодотворении накладывает икру на кожу спины самки; кожа, разрастаясь, образует ячейки вокруг яиц. В этих ячейках детеныши проходят все свое развитие и выходят на свободу по окончании превращения уже в совершенно развитой форме. У спиномешочницы (Notodelphys orifera, табл. II, рис. 2) самка имеет на спине сумку, в которой развиваются яйца, а вылупившиеся из них детеныши подвергаются превращению. У большей же части Б. гадов икра развивается в воде, и детеныши не пользуются никаким попечением со стороны родителей. Зародыш развивается так быстро, что детеныши вылупляются из яйца уже через несколько дней, на весьма низкой ступени развития. Откладывается ли икра шнурками или комками, отдельные яички всегда бывают окружены клейким, разбухающим в воде слизистым веществом, которое играет роль защитительного покрова от вредного влияния воды на яйцо. Желток в той половине, которая обращена кверху, бывает более темного цвета, что зависит от черно-коричневого пигмента, находящегося на его периферии. Этот более темный желток есть образовательный желток и из него только и развивается зародыш, а светлый желток служит для питания. В этой темной половине начинается процесс дробления (табл. III, рис. 20), который и идет здесь быстрее, чем на светлой половине, на которой сегментационные шары бывают крупнее и не так многочисленны. По окончании дробления внутри образовавшейся клеточной массы возникает полость, которая лежит ближе к верхней половине, чем к нижней, вследствие чего последняя имеет больший удельный вес. На верхней половине образуется зародыш с первичной бороздкой и спинными валиками; он быстро обрастает желток, еще до того, как спинные валики сомкнутся в мозговую трубочку. После образования жаберных возвышений, но еще до появления ротового отверстия снабженные коротким хвостом зародыши (головастики), представляющие различную степень развития, смотря по виду, покидают свои яйцевые покровы и прикрепляются к слизистым остаткам икры с помощью двух присасывательных ямок (табл. III, рис. 21 и 22). Раньше прочих вылупляются личинки некоторых жаб — еще прежде, чем покажутся следы наружных жабр на жаберных возвышениях, отделенных друг от друга щелями; но большая часть Б. гадов оставляет яйцо уже с более или менее развитыми тремя парами наружных жабр, которые быстро разрастаются. Только большие личинки повитушки (Alytes) выходят из яиц, уже пройдя стадию с наружными жабрами. За исключением этих последних, головастики сначала не способны сами принимать пищу и питаются запасами желтка, оставшимися в кишечном канале. Затем тело удлиняется, хвост получает значительную длину, прежде едва заметные глазные точки яснее проявляются под кожей головы, рот открывается; движения головастика становятся ловчее и смелее, и он уже начинает отыскивать пищу самостоятельно. Вскоре наружные жабры исчезают, кожа обрастает жаберные щели, из которые остается только одна, через которую вытекает вода из обеих жаберных полостей; на место наружных жабр развиваются внутренние жабры, именно на боковой стенке всех 4 жаберных дуг образуются гребенчатые жаберные листочки (табл. III, рис. 27 и 28а). Губы получают роговые чехлики, и таким образом возникает роговой клюв, служащий для обтачивания листьев и других пищевых веществ. Пищеварительный канал значительно удлиняется и образует улиткообразные извивы (табл. III, рис. 26), из глотки выпячиваются два легочных мешка (табл. III, рис. 29), которые вместе со внутренними жабрами служат для дыхания: по временам головастики поднимаются на поверхность воды, чтобы вдохнуть в себя воздух. При дальнейшем развитии на теле головастика, на самой границе хвоста, появляются задние конечности в виде маленьких зачаточных придатков; жаберный аппарат постепенно уступает свое место легким и затем наступает линька, после которой жаберные листики исчезают и появляются передние конечности, уже давно развившиеся под кожей, исчезает роговой клюв и большие глаза выступают наружу. Таким образом головастик превращается в дышащее воздухом четвероногое животное, снабженное коротким хвостом; хвост постепенно укорачивается и скоро обращается в маленький отросток; молодые животные оставляют воду и выходят на сушу. Элементы хвоста уничтожаются выступающими из крови блуждающими клетками (фагоцитами). По исчезновении последних остатков хвоста детеныши получают окончательную форму. Так происходит развитие огромного большинства Б. Г., но известно одно очень интересное исключение, именно у кокки, или мартиникской квакши (H ylodes martinicensis), из яиц выходят не головастики, а вполне сформировавшиеся молодые квакши, отличающиеся от взрослых только присутствием очень маленького хвоста, который скоро исчезает.
Время, в течение которого совершается превращение, очень различно и находится в зависимости, с одной стороны, от вида, а с другой — от климата и погоды. Большей частью продолжительность развития и величина личинок находятся в прямой зависимости, т. е. чем медленнее идет развитие, тем большей величины достигают личинки; так, жабы развиваются быстрее прочих Б. Г. и вместе с тем имеют наименьших личинок; из европейских видов чесночницы (Pelobates) имеют самых крупных головастиков, и для ее превращения требуется почти вдвое более времени, чем для зеленой лягушки (Rana escule n ta, табл. I, рис. 2), и вчетверо, чем для жабы вонючей (Bufo calamita). Высокая температура ускоряет метаморфоз.
Б. гады — земноводные животные в полном значении этого слова, так как в молодости, когда имеют наружные или внутренние жабры, они живут в воде, а во взрослом состоянии, когда разовьются легкие и исчезнут жабры, держатся на суше, входя в воду только для размножения (напр., жабы) или же проводят часть жизни в воде, а часть на суше (напр., лягушки). Живущие во взрослом состоянии постоянно на суше особенно любят сырые и темные места и отличаются тем, что плавательные перепонки на их задних ногах или совсем не развиты (напр., у мартиникской квакши Hylodes martinicensis) или же развиты не вполне (напр., у наших жаб), а лишь у весьма немногих не вполне развиты (например, у чесночницы, Pelobates); напротив, у видов, живущих как на суше, так и в воде, задние ноги обыкновенно снабжены полными плавательными перепонками. Б. Г. — преимущественно ночные животные, хотя многие из них и днем обнаруживают весьма оживленную деятельность. Подвижностью они превосходят других земноводных: довольно хорошо ходят, прыгают необыкновенно ловко и сравнительно большими скачками, превосходно плавают и ныряют; некоторые (квакша, табл. I, рис. 10) лазят по деревьям и кустарникам при помощи присасывательных кружочков на концах своих пальцев и даже перепархивают с ветки на ветку (летучая лягушка, Khacophorus Reinwardtii, табл. I, рис. 9), причем роль летательного аппарата играют чрезвычайно широкие плавательные перепонки задних ног; иные могут вырывать для себя норы (чесночница, Pelobates; повитушка, Alytes). За весьма немногими исключениями Б. Г. — общественные животные, так как всегда селятся в очень большом числе в одной и той же местности; общественность эта чисто внешняя, так как каждая особь живет сама по себе и вовсе не заботится об остальных. Чувства их развиты лучше, чем у остальных земноводных: они имеют развитое зрение, тонкий слух и одарены острым обонянием, обладают осязанием и одарены, хотя и в слабой степени, вкусом. Б. Г. обнаруживают: способность распознавать местность, память, сообразительность, умеют пользоваться опытом, осторожны и робки относительно других животных и даже до известной степени хитры, когда дело идет о том, чтоб избежать опасности или завладеть добычей. Питаются Б. Г. преимущественно насекомыми, червями, улитками, иногда же им пищей служат икра и маленькие рыбки; самые крупные виды этого отряда пожирают даже маленьких млекопитающих и птиц (напр., американская воловья лягушка, Rana mugiens); как настоящие хищники, они едят только живую добычу, пойманную ими самими. Головастики употребляют преимущественно растительную пищу, но не ограничиваются ею, как это прежде думали, а едят также мелких животных и животные вещества. В противоположность головастикам и большей части других земноводных Б. Г. во взрослом состоянии обладают голосом иногда настолько громким, что он бывает слышен на несколько верст в окружности (напр., воловья лягушка). Особенно громок голос у самцов, снабженных голосовыми мешками, играющими роль резонаторов. Б. Г., подобно всем земноводным, нуждаются в тепле и не переносят холода. При наступлении холодного времени года они уходят в норы, под корни и в дупла деревьев или глубоко зарываются в ил и там впадают в зимнюю спячку, продолжающуюся до возвращения теплого времени. Во время спячки холод на них не действует: они могут промерзать насквозь так, что во внутренних органах, напр., в сердце, заключается лед, и тем не менее весной оказываются и продолжают жить. Весеннее пробуждение наступает у различных видов различно: в то время как одни пробуждаются ранней весной, когда еще снег и лед не сошли с полей, другие показываются из своих убежищ только в конце апреля и в начале мая. Если летом во время сильной засухи пересохнут все стоячие воды и жившие в них Б. Г. не найдут возможности перебраться в другое место, более богатое водой, то они впадают в более или менее продолжительную летнюю спячку, которая оканчивается с первым сильным дождем, наполняющим их жилища водой. В таких случаях на тех местах, где во время засухи не было видно ни одного гада, вдруг появляется целая масса их. Это, вероятно, и послужило поводом к рассказам о "лягушечьих дождях". Впрочем, эти рассказы могут иметь и другой источник. Возможно, что масса развившихся из головастиков и вышедших на сушу молодых животных подхватывается порывом бури, который проносит из через известное пространство и затем роняет на землю в другом месте.
По окончании превращения дальнейший рост Б. Г. идет очень медленно. Способность размножаться проявляется несколько лет спустя. Рост Б. Г., т. е. увеличение массы тела, по-видимому, продолжается до глубокой старости. По крайней мере известно, что жабы, содержавшиеся в неволе десять лет и все время получавшие обильный и хороший корм, далеко не достигали такой величины, какая встречается у некоторых особей, живущих на свободе. Ввиду медленного развития и сравнительно малой подвижности этих животных можно допустить, что жизнепродолжительность их очень велика. Средняя жизнепродолжительность Б. Г. увеличивается еще их живучестью; они продолжают жить даже при таких обстоятельствах, при которых смерть была бы неизбежна для большинства других животных. Так, замуравленные жабы оставались живыми в течение целых месяцев, не получая никакой пищи; лягушка со вскрытым позвоночным мозгом продолжает бодро прыгать; из желудка убитых змей иногда удавалось извлекать проглоченных ими жаб, которые были еще живы, несмотря на то, что задние ноги их уже начали перевариваться. Однако живучесть эта не безгранична и в некоторых случаях Б. Г. умирают так же скоро, как и другие позвоночные животные; в особенности все они с большим трудом переносят недостаток влажности.
По отношению к человеку Б. Г. являются преимущественно полезными животными. Причиняемый ими вред весьма незначителен и ограничивается тем, что они иногда поедают полезных для человека насекомых, а некоторые более крупные виды (напр., бурая и воловья лягушки) поедают икру мелких рыб, а иные (Воловья лягушка) иногда нападают на молодых домашних птиц (напр., утят). Приносимая Б. Г. польза несравненно значительнее и с избытком покрывает причиняемый ими вред. Именно, эти земноводные полезны как деятельные истребители вредных для человека насекомых и улиток, которых они пожирают в огромном количестве; польза эта настолько ощутительна, что многие благоразумные садоводы покровительствуют Б. Г., особенно жабам, а англичане даже покупают последних для того, чтобы очищать свои сады и огороды от различных вредных насекомых и улиток. Кроме того, задние ноги зеленой лягушки (Rana esculenta) употребляются в Западной Европе, особенно во Франции, в пищу и считаются лакомством. Наконец, лягушки и некоторые другие Б. Г. служили и служат важным материалом для физиологических, гистологических и эмбриологических работ и первые стали необходимой принадлежностью каждой физиологической лаборатории. Кроме человека, Бесхвостые гады имеют важное значение для многих животных, составляя их исключительную пищу или же более или менее важную составную часть последней. К числу таких животных принадлежат болотные и многие хищные птицы, змеи, хищные рыбы, а также и крупные водяные хищные насекомые, поедающие их икру и головастиков.
Средства защиты земноводных от преследования многочисленных врагов их очень ограничены. Лишь весьма немногие Б. гады имеют сильно развитые кожные железы, выделяющие ядовитый и едкий сок, который делает их несъедобными и охраняет от нападений. Так, у жаб железы спины и околоушные выделяют похожую на молоко жидкость; жидкость эта слегка желтоватого цвета, горького вкуса, едка и имеет неприятный запах; она представляет кислую реакцию и на воздухе свертывается. Опыты показали, что действие ее не уничтожается даже от высушивания. Будучи привита птицам, она убивает их в несколько минут, мелких млекопитающих (морских свинок) и собак — не более как в полтора часа. На жаб этот яд не действует, хотя убивает лягушек. Яд жаб служит им лишь средством для защиты, но не для нападения, так как эти животные не имеют таких органов, которые давали бы им возможность вводить его в кровь животных. Для человека жабы совершенно безвредны, так как яд их на человеческую кожу не действует и, только попав на более нежные места, напр. на глаза, вызывает довольно сильную боль. Другие, напр. лягушки, могут выпрыскивать на врага воду или мочу из клоаки, но это средство защиты мало надежно — оно может помочь только в том случае, когда преследователь испугается внезапного душа. Некоторые обладают охранительной окраской; так, квакша (Hyla arborea), живущая между листьями деревьев и кустарников, бывает зеленого цвета; земляная жаба (Bufo vulgaris) отличается землисто-бурым или грязно-серым цветом, сливающимся с окружающей ее почвой, по которой она ползает; илисто-серая сверху жерлянка (Bombinator igneus, табл. I, рис. 7) совершенно подходит по окраске к болотному илу, на котором живет; зеленая лягушка (Rana esculenta), держащаяся обыкновенно в траве, имеет сверху серо-зеленый цвет. Многие Б. Г. спасаются от преследования врагов только тем, что ведут очень скрытый образ жизни, прячась в трудно доступных местах, из которых выходят только с наступлением сумерек.
Как мы уже видели, окраска Б. гадов довольно разнообразна; вообще в этом отряде нередки ярко окрашенные представители, особенно между обитателями жарких стран. Цвет этих видов бывает различен не только у разных видов, но у некоторых замечаются также и половые различия в окраске; так, у обыкновенной жабы (Bufo vulgaris) самец имеет одноцветное светло-серое брюхо, а у самки на светло-сером брюхе расположено много темных пятен; у квакши (Hyla arborea) самец имеет черновато-бурое, а самка беловатое горло. Наконец, у некоторых окраска изменяется соответственно с окраской окружающей среды, напряженностью дневного света и проч. (напр., у квакши, у зеленой жабы). Изменения эти зависят от того, что в коже таких животных красящие вещества (пигменты) различного цвета заключаются в так называемых хроматофорах, т. е. клетках, способных расширяться и сокращаться под влиянием нервной системы; преобладающим является тот цвет, соответственные хроматофоры которого в данный момент расширены; при расширении хроматофоров другого цвета происходит изменение окраски.
Б. гады распространены, за исключением Крайнего Севера, по всей земле; преимущественно же теплые климаты богаты большими и разнообразно окрашенными видами; всего богаче этими животными тропическая область (Южная Америка); благодаря более совершенным органам передвижения Б. гады имеют более широкое распространение, чем прочие земноводные, но и у них распространение отдельных видов ограничивается естественными препятствиями: морями, горами и пустынями. Наиболее широкое распространение имеют роды Hyla, Bufo и Rana. Б. гады, по-видимому, появились на земле после других земноводных (хвостатых гадов); по крайней мере самые древние остатки их найдены в верхних эоценовых пластах. Собственно лягушки (Rani d ae) найдены в пресноводных пластах различных местностей Германии, Франции и около Бомбея в Индии. Остатки родов Asphaerios и Palaeobatrachus, из Богемии и Зинбенгебирга, очень близки к лягушкам; в Энингене, в Швейцарии, найдены остатки рода Palaeophryne очень близкого к современным жабам.
Судя по истории развития, необходимо допустить, что Б. гады происходят прямо от хвостатых, а через них косвенно от рыб, и именно от твердочешуйных (осетровых рыб), судя по сходству строения различных внутренних органов у тех и других. В самом деле, поздние стадии головастиков по своему строению совершенно соответствуют хвостатым гадам (тритон, саламандра), сначала имеющим наружные, а потом — тем, у которых бывают внутренние жабры. Более ранние стадии головастиков соответствуют рыбам, и у некоторых форм сходство головастиков с рыбами выражается не только во внутреннем строении, но и во внешней форме (напр. у личинки Dactylethra capensis, табл. II, рис. 1в.).
Ловля Б. гадов не затруднительна; они настолько малоподвижны, что их можно ловить руками или же сачком. Несколько более затруднительно отыскивание их, так как эти животные очень пугливы и обыкновенно днем прячутся, так что удачное отыскивание Б. гадов зависит от знакомства с их образом жизни и нравами. Пойманных животных помещают в террарии и кормят дождевыми червями и различными насекомыми (мухами, мучными червями, тараканами и прочими). Когда настает время размножения, то в террарии помещают большие сосуды с водой с таким расчетом, чтобы для каждой пары было по сосуду. Когда икра отложена, то ее перемещают в другой сосуд из предосторожности, чтобы взрослые животные своими движениями не причинили бы ей вреда. Затем икру надо оберегать от толчков и сотрясений и ежедневно менять воду в занимаемом ею сосуде, причем самую перемену воды надо производить как можно осторожнее. Испортившиеся икринки необходимо тщательно удалять. Вылупившихся головастиков кормят растертыми облатками, растительными веществами, очень маленькими кусочками мяса и т. п. По окончании метаморфоза в первое время им надо давать самых мелких червей и насекомых, так как с более крупными они еще не в силах справиться.
Набивка Б. гадов производится таким образом: делают разрез кожи около рта, и затем кожа снимается с тела, как перчатка; снятая кожа насыпается нагретым песком, и полученный таким образом препарат высушивается. Однако такое препарирование требует большого навыка и умения, в противном случае препараты бывают очень плохи, так как животное утрачивает свою форму. Поэтому гораздо предпочтительнее сохранять Б. гадов в консервирующих жидкостях. Для этой цели чаще всего употребляется винный спирт, разбавленный водой (2 части спирта и 1 часть воды для взрослых животных и поровну того и другого для головастиков); для сохранения цвета к спирту прибавляют квасцы (15 г на 1 кг), которые предварительно растворяют в воде, прибавляемой к спирту. Головастиков сохраняют также в глицерине.
Б. гады делятся на три группы и на шесть семейств. I группа: дископалые (Discodactylia), имеют язык; пальцы широкие, с расширенными подушечками на концах. 1 семейство: древесницы (Hylidae). Сюда относятся: квакша (Hyla arborea, табл. I, рис. 10); спиномешочница (Notodelphys ovifera, табл. I, рис. 2); мешконоска (Nototrema marsupiatum, табл. I, рис. 11); летучая квакша (Rhacophorus Reinwardtii, табл. I, рис. 9); кокки (Hylodes marticensis) и пр. II группа: остропалые (Oxydactylia), с подвижным языком и острыми концами пальцев. 2 семейство: лягушки (Ranidae). Сюда относятся: зеленая лягушка (Rana esculenta, табл. I, рис. 2); бурая лягушка (R. temporaria, табл. I, рис. 3); воловья лягушка (R. mugiens); рогатая лягушка (Ceratophrys cornutus, табл. I, рис. 6); ильная лягушка (Pelodytes punctatus) и пр. 3 семейство: чесночницы (Pelobatidae). Сюда относятся: чесночница бурая (Pelobates fuscus); повитушка (Alytes obstetricans, табл. I, рис. 5); жерлянка (Bombinator igneus, табл. I, рис. 7) и друг. 4 семейство: жабы (Bufonidae). К этому семейству принадлежат: жаба обыкновенная (Bufo vulgaris, табл. I, рис. 8); жаба зеленая (B. variabilis), жаба вонючая ((B. calamita), носачка (Rhynophrynus dorsalis) и проч. III группа: безъязычные (Aglossa). Языка нет, пальцы оканчиваются остриями или когтями. 5 семейство: пипы (Pipidae); сюда относится: пипа суринамская (Pipa dorsigera, табл. I, рис. 1). 6 семейство: коготницы (Dactylethridae). Сюда принадлежит наперсточница капская (Dactylethra capensis, табл. I, рис. 1).
Бесхвостые гады принадлежат к числу животных, о которых существует масса различных суеверных и вздорных рассказов, главнейшим источником которых является незнание жизни этих существ. Так, например, многие убеждены, что не только жабы, но и лягушки очень ядовиты; рассказывают, что будто бы лягушки и жабы бросаются на преследующего их человека и впиваются в него, будто жабы по ночам высасывают молоко у коров, что от присутствия лягушек вода становится светлее и холоднее, будто, если убить лягушку, то непременно будет дождь, и т. д.
Важнейшие сочинения о Б. гадах следующие: а) общего содержания: Hoffmann, "Amphibien" в Bronn's "Klassen und Ordnungen des Thierreichs" (т. 17, отд. 2, Лейпциг и Гейдельберг, 1879—80); Knauer, "Naturgeschichte der Lurche. Amphibiologie" (2 изд., Вена, 1883); b) по систематике: Roesel von Rosenhof, "Historia naturalis ranarum nostratium" (Нюрнберг, 1658); Daudin, "Histoire nat urelle des Rainetes, Grenouilles et des Crapauds" (Париж, 1882); Günther, "Catalogue of the Batrachia salientia in the collection of the British Museum" (Лондон, 1858); Bruch, "Beiträge zur Naturgeschichte und Classifikation der nackten Amphibien" ("Würzburg. naturg. Zeitsch.", 1862); Mivart, "On the classification of the Anurous Batrachians" ("Proceed. zool. Soc.", 1869); Steindachner, "Amphibien. Reise der Fregatte Novara" (Вена, 1867); его же, "On the families of the raniform Anura" ("Journ. of the Acad. of Philadelphia", 1866 и 1867); с) по анатомии: Dugés, "Recherches sur l'osté ologie et la myologie des Batraciens" (Париж, 1835); Ecker und Wiedersheim, "Die Anatomie des Frosches" (Брауншвейг, 1869—82); d) по истории развития: C. Vogt, "Untersuchungen üb er die Entwickelungsgeschichte der Geburtshelferkroete" (Золотурн, 1842); Rusconi, "Dé veloppement de la grenouille commune" (Милан, 1826); van Bambeke, "Recherches sur le développement de Pélobate brun" ("Mé m. cour. Acad. de Belg." (т. 34, 1868); Goette, "Entwickelungsgeschichte der Unke" (Лейпциг, 1874); Martin Saint-Ange, "Recherches anatomiques et phisiologiques sur les organes transitoires et la mé tamorphose des Batraciens" ("Ann. Sc. nat.", т. 24, 1831).
|
Page was updated:Tuesday, 11-Sep-2012 18:14:41 MSK |