| [ начало ] | [ Г ] |
Головной мозг
(Encephalon). Содержание: А. Анатомия головного мозга человека: 1) строение Г. мозга, 2) оболочки мозга, 3) кровообращение в Г. мозгу, 4) ткань мозга, 5) ход волокон в мозгу, 6) вес мозга. — В. Эмбриональное развитие Г. мозга у позвоночных животных. — С. Строение Г. мозга в различных классах позвоночных животных. — D. Функции Г. мозга (физиол.): 1) продолговатый мозг, 2) мозговые ножки, 3) мозжечок, 4) четырехолмия, 5) полосатые тела, 6) полушария мозга.
A. Г. мозг человека. 1) Строение Г. мозга. Черепная часть центральной нервной системы человека заключает в себе следующие большие отделы: большой мозг (cerebrum), малый, или мозжечок (cerebellum), и продолговатый мозг (medulla oblongata).
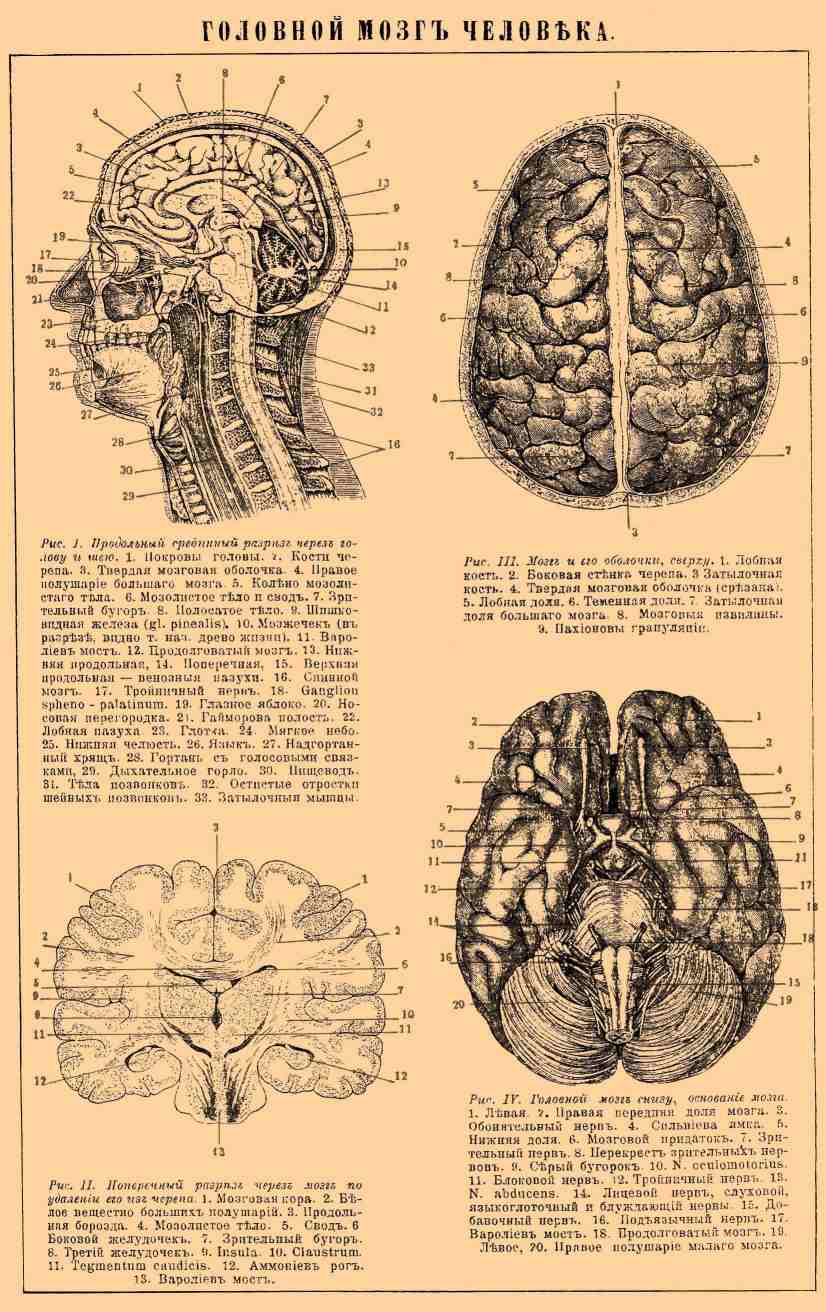
Рис. 1. Продольный срединный разрез через голову и шею. 1. Покровы головы. 2. Кости черепа. 3. Твердая мозговая оболочка. 4. Правое полушарие большого мозга. 5. Колено мозолистого тела. 6. Мозолистое тело и свод. 7. Зрительный бугор. 8. Полосатое тело. 9. Шишковидная железа (gl. pinealis). 10. Мозжечок (в разрезе, видно так называемое древо жизни). 11. Варолиев мост. 12. Продолговатый мозг. 13. Нижняя продольная, 14. Поперечная, 15. Верхняя продольная — венозные пазухи. 16. Спинной мозг. 17. Тройничный нерв. 18. Ganglion spheno-palatinum. 19. Глазное яблоко. 20. Носовая перегородка. 21. Гайморова полость. 22. Лобная пазуха. 23. Глотка. 24. Мягкое нёбо. 25. Нижняя челюсть. 26. Язык. 27. Надгортанный хрящ. 28. Гортань с голосовыми связками. 29. Дыхательное горло. 30. Пищевод. 31. Тела позвонков. 32. Остистые отростки шейных позвонков. 33. Затылочные мышцы.
Рис. II. Поперечный разрез через мозг по удалении его из черепа. 1. Мозговая кора. 2. Белое вещество больших полушарий. 3. Продольная борозда. 4. Мозолистое тело. 5. Свод. 6 Боковой желудочек. 7. Зрительный бугор. 8. Третий желудочек. 9. Insula. 10. Claustrum. 11. Tegmentum caudicis. 12. Аммониев рог. 13. Варолиев мост.
Рис. III. Мозг и его оболочки, сверху. 1. Лобная кость. 2. Боковая стенка черепа. 3. Затылочная кость. 4. Твердая мозговая оболочка (срезана). 5. Лобная доля. 6. Теменная доля. 7. Затылочная доля большого мозга. 8. Мозговые извилины. 9. Пахионовы грануляции.
Рис. IV. Головной мозг снизу, основание мозга . 1. Левая, 2. Правая передняя доля мозга. 3. Обонятельный нерв. 4. Сильвиева ямка. 5. Нижняя доля. 6. Мозговой придаток. 7. Зрительный нерв. 8. Перекрест зрительных нервов. 9. Серый бугорок. 10. N. oculomotorius. 11. Блоковой нерв. 12. Тройничный нерв. 13. N. abducens. 14. Лицевой нерв, слуховой, языкоглоточный и блуждающий нервы. 15. Добавочный нерв. 16. Подъязычный нерв. 17. Варолиев мост. 18. Продолговатый мозг. 19. Левое, 20. Правое полушарие малого мозга.
Уже невооруженным глазом в нем различаются два вещества — серое и белое, которые оказываются неодинаковыми и по микроскопическому строению, и по своей роли в отправлениях Г. мозга. Серое вещество то образует сплошные слои на поверхности мозга (корка большого и малого мозга); то выстилает его полости, наз. желудочками; то, наконец, скопляется внутри белого вещества в виде гнезд, называемых узлами, ядрами, центрами и пр. Большой мозг представляет собою почти половину шара, несколько сплющенную с боков; его плоская поверхность обращена вниз и наз. основанием мозга (basis cerebri). Глубокая щель (fissura pallii) делит его на правое и левое полушария (hemispherae cerebri); последние в средней части, где щель менее глубока и не доходит до основания мозга, соединяются между собою при помощи мозолистого тела. Мозолистое тело, или большая спайка (corpus callosum, comissura magna), имеет форму дуги, которой передний конец, наз. головкой, загибается вниз и доходит до основания мозга, а задний, утолщенный, висит над четверохолмием; оно состоит из пучков белого вещества, которые имеют поперечное направление и из мозолистого тела продолжаются в белое вещество полушарий, служа здесь крышкой боковых желудочков. Поверхность полушарий большого мозга изрезана многочисленными бороздами (fissurae, sulci); одни из них существуют на всяком мозгу и отличаются постоянством очертаний, другие представляют значительные вариации в развитии и могут совсем отсутствовать. Типические борозды, помимо постоянства, отличаются большею глубиною (3,5-4,7 см) и ранним появлением (между 3-7 месяцем утробной жизни). Борозды разбивают поверхность полушарий большого мозга на извилистые полоски - валики (извилины — gyri); несколько соседних извилин составляют долю полушария. В каждом полушарии большого мозга различают четыре доли. Лобная, или передняя, доля (lobus frontalis anterior) нижнею своею поверхностью лежит в передней черепной яме, а верхнею и боковыми поверхностями прикасается к лобной и отчасти теменным костям. Теменная, или верхняя, доля (lobus parietalis, superior) прилежит к теменной кости; затылочная, или задняя (lobus occipitalis, рostеriоr) — к задней части теменной и к затылочной кости до верхней дугообразной ее линии; височная, или нижняя (I. temporalis, inferior) лежит в средней черепной яме. Границами между этими долями отчасти служат некоторые из типических борозд, отчасти условно проводимыe линии; так, глубокая борозда на нижней поверхности полушария, наз. Сильвиевой ямой (fossa Sylvii), отделяет здесь лобную долю от височной; борозда на наружной (выпуклой) поверхности полушария, почти вертикально поднимающаяся несколько впереди середины последнего и наз. центральной, или Роландовой, отграничивает сзади лобную долю от теменной; горизонтальная ветвь вилообразной борозды, служащей продолжением Сильвиевой ямы на наружную поверхность полушария и наз. Сильвиевой бороздой (fissura Sylvii), отделяет височную долю от вышележащих (лобной и теменной). Сильвиева борозда отличается своею глубиною, и дно ее закрыто сходящимися краями, образуя скрытую, или центральную, долю полушария (lobulus opertus, l. centralis). Если у новорожденного младенца центральная долька видна с поверхности, это указывает на недоразвитость мозга. Каждая мозговая доля покрыта определенным числом извилин; на лобной доле лежат gyrus centralis anterior, g. frontalis primus, secundus et tertius; три извилины лежат на теменной, три на наружной поверхности височной доли. Нижняя поверхность полушарий и внутренняя поверхность их (т. е. та сторона, которой они обращены друг к другу, до соединения их посредством мозолистого тела) также имеют свои типические борозды и извилины. Кроме того, есть множество мелких извилин, отличающихся чрезвычайными индивидуальными различиями и образующиеся позднее типических (после 7-го месяца утробной жизни). Под нижней поверхностью затылочных долей большого мозга лежит мозжечок, а под нижней поверхностью этого последнего — продолговатый мозг. Последний есть непосредственное продолжение спинного мозга, отчего его наз. также луковицей спинного мозга (bulbus medullae spinalis); границей между продолговатым и спинным мозгом считают уровень затылочной дыры — место отхождения первой пары шейных нервов. Продолговатый мозг — конической формы тело, обращенное суженною своею частью вниз и лежащее на основной части затылочной кости. На передней и задней поверхностях продолговатого мозга проходит по бороздке, которые суть продолжения передней и задней продольных борозд спинного мозга. Нервные волокна, составляющие продолговатый мозг, собираются в несколько пучков. По обеим сторонам передней продольной борозды продолговатого мозга лежит по пучку, наз. пирамидами. Если последние раздвинуть, то видно, что часть волокон левой пирамиды переходит в правую пирамиду, а часть волокон правой переходит в левую; это место наз. перекрестом пирамид (decussatio pyramidum). Кнаружи от обеих пирамид лежит по бугру овальной формы, наз. оливами (olivae), а рядом с последними — по пучку волокон, наз. веревчатыми телами (corpora restiformia), которые в нижней своей части отделены друг от друга задней продольной бороздой, а вверху расходятся под острым углом, образуя задний угол, так наз. ромбовидной ямки, и вступают в мозжечок под назв. ножек мозжечка к продолговатому мозгу (crura cerebelli ad medullam oblongatam ). В углу между расходящимися веревчатыми телами натянута весьма тонкая пластинка треугольной формы, образующая часть крыши 4-го желудочка и называемая задним мозговым парусом (velum medullare posticum). Каждое веревчатое тело в верхней своей части состоит из двух пучков, клиновидного (funiculus cuneatus), образующего на верхнем конце клиновидный бугорок (tuberculum cuneatum), и тонкого (funiculus gracilis), с утолщением, называемым булавой (clava). Продолговатый мозг — белого цвета. Верхняя (задняя) его поверхность, служащая дном полости, назыв. 4 желудочком, покрыта слоем серого вещества, который есть продолжение серого вещества спинного мозга. Далее скопления серого вещества находятся также в веревчатых телах и в оливах. Впереди продолговатого мозга лежит Варолиев мост (pons Varolii). Он представляет толстую и широкую полосу белого мозгового вещества, которая огибает передний конец продолговатого мозга, соединяясь с ним. Боковые его части, соединяющие Варол. мост с мозжечком, называются ножками мозжечка к мосту (crura cerebelli ad pontem). Из переднего края Варол. мозга выходит с правой и левой стороны по широкой и толстой пластинке мозговой ткани, направляющиеся к полушариям большого мозга и вступают в них; эти пластинки называются ножками мозга (pedunculi, crura, caudex cerebri). Нижняя, более широкая часть ножки называется основанием ее (basis pedunculi), a верхняя, более толстая, называется покрышкой ножки (tegmentum caudicis). Эти два белых пучка отделены друг от друга слоем серого вещества. Впереди от Варол. моста лежит пластинка серого вещества, через которую кровеносные сосуды вступают в ткань мозга; это — задняя продырявленная пластинка (lamina perforata posterior), впереди которой лежат рядом два бугорка белого вещества — сосковидные тела (corpora mammilaria); в центральной их части есть небольшое скопление серого вещества. Тотчас впереди последних помещается серый бугор (tuber cinereus); от верхушки его отходит вниз тонкий отросток, называемый воронкой (infundibu lum), на котором висит тело овальной формы, называемое придатком (hypophysis cerebri), наполовину состоящее из нервной ткани; нижняя же (передняя) доля придатка состоит из соединительной ткани. На обоих полушариях большого мозга из-под его ножек выходит по полоске (tractus opticus) нервной ткани, которые идут вперед, приближаясь друг к другу; впереди серого бугра они образуют перекрест (chiasma, decussatio nervorum opticorum), т. е. волокна правой полоски переходят на основание левого полушария, а волокна левой полоски на правое полушарие, и затем опять расходятся в виде шнурков, называемых зрительными нервами (nervi орtiсi). Наконец, впереди и под зрительным перекрестом лежит пластинка серого вещества, соединяющая оба полушария (lamina cinerea terminalis); боковые ее треугольные части называются передними продырявленными пластинками (lamina, substantia perforata anterior). Мозжечок состоит из двух полушарий (hemispherae cerebelli), связанных между собою узкою срединною частью, червячком (vermis superior et inferior). На обеих поверхностях мозжечка проходят многочисленные борозды, отличающиеся своим однообразным направлением: почти все они параллельны заднему краю мозжечка, и очень многие параллельны друг другу. Глубина этих борозд не превышает 2 1/2 см. Полушария мозжечка соединены с Варолиев. мостом, с продолговатым мозгом и с четверохолмием посредством пучков нервных волокон, называемых ножками; о ножках к Варол. мосту и продолговатому мозгу уже было сказано. Ножки к четверохолмию (crurа cerebelli ad corpus quadrigeminum) лежат под ножками к Варолиеву м., идут прямо вперед и исчезают под четверохолмием; пластинка, натянутая между этими ножками и составляющая часть крышки 4-го желудочка, наз. передним мозговым парусом (velum medullare anticum). Мозжечок по поверхности покрыт слоем серого вещества, образующим корку мозжечка. Под наружным серым слоем скрывается скопление белого вещества, называемого древом жизни (arbor vitae). В разрезе полушария мозжечка видно, что от центрального белого вещества отходят несколько ветвей белого цвета; от этих ветвей отходят новые ветви и т. д. Вокруг этих разветвлений белого вещества поверхностное серое расположение как бы в виде листьев. Разрез червячка представляет такую же картину. Кроме поверхностного серого вещества, последнее встречается среди центрального белого в виде серых узлов. Между мозжечком и лежащим под ним продолговатым мозгом помещается полость, наз. 4-м мозговым желудочком; дном последнего служит четырехугольное углубление на верхней (задней) поверхности продолговатого мозга. Берега этого углубления, наз. ромбовидной ямкой (fossa rhomboidea), образуются спереди ножками мозжечка к четверохолмию, а сзади — веревчатыми телами. Верхнюю стенку-крышку желудочка образуют мозговые парусы. Над парусами лежит отросток мягкой мозговой оболочки, по богатству сосудами наз. нижним сосудистым сплетением. В переднем парусе есть отверстие (hiatus Magendii), сообщающее полость 4-го желудочка с подпаутинным пространством (см. ниже). Четвертый желудочек (ventriculus cerebri quartus) — есть продолжение центрального канала спинного мозга; сообщением между ними служит отверстие в заднем углу ромбовидной ямки; слой серого вещества, выстилающий последнюю, есть продолжение серого вещества спинного мозга. От переднего угла ромбов. ямки к заднему идет бороздка, берега которой образуют два продольных валика, наз. круглыми канатиками (funiculi teretes). Задний угол ромбов, ямки назыв. писчим пером (calamus scriptorius); два углубления темного цвета в последнем наз. серыми крыльями (alae cinereae). В передней части ромбов. ямки лежат два серых бугорка (locus coeruleus). Белые поперечные полоски в средней части ромбов. ямки назыв. слуховыми (striae, chordae acusticae); в переднем углу ямки лежит отверстие, ведущее в Сильвиев водопровод (aquaeductus Sylvii). Последний есть узкий канал в массе четверохолмия, выстланный слоем серого вещества и соединяющий четвертый мозговой желудок с 3-м. Третий желудочек лежит под средней частью мозолистого тела, а по бокам последнего помещаются правый и левый боковые желудочки. Каждый боковой, или трехрогий, желудочек (ventriculus lateralis tricornis) состоит из средней части (cella media) и трех рогов, т. е. дугообразных продолжений. В переднем роге, направляющемся к лобной доле, лежит тело грушевидной формы, верхний слой которого состоит из сплошного серого вещества, а в более глубоких слоях полоски серого цвета чередуются с полосками белого — это хвостатое тело (corpus caudatum), составляющее часть полосатого тела (corpus striatum). Позади полосатого тела находится так наз. зрительный бугор (thalamus opticus), большая часть которого принадлежит третьему желудочку. Между зрительным бугром и полосатым телом тянется полоска сероватого цвета, доходящая до заднего рога и называемая роговой полосой (stria cornea). Возвышение на внутренней поверхности заднего рога, направляющегося к затылочной доле полушария, наз. птичьей шпорой или малой ногой морского коня (calcar avis, pes hippocampi minor); возвышение на наружной стенке того же рога — eminentia collateralis Meckelii. На дне нижнего рога, лежащего в височной доле, проходит возвышение, наз. большой ногой морского коня или Аммониевым рогом (pes hippocampi major, cornu Ammonis). Каждый из зрительных бугров, составляющих боковые стенки узкой вертикальной щели, наз. третьим мозговым желудочком, представляет собою трехгранной формы тело с закругленными краями, верхняя поверхность которого покрыта весьма тонким слоем белого вещества (Stratum zonale); сам же зрительный бугор состоит из темно-серого вещества. Задняя часть бугра, наз. подушкой (pulvinar), отделяется неглубокой бороздкой от передней, наз. передним бугорком; возвышение на подушке наз. передним (наружным) коленчатым телом. От задней поверхности зрительного бугра отходит толстая полоса, направляющаяся к основанию мозга под именем зрительного пути (tractus opticus). Над третьим желудочком тянется полоса нервной ткани, прилегающая к нижней поверхности средней части мозолистого тела. Эта полоса, наз. сводом (fornix), делится кпереди и кзади на две передние и задние ножки свода. Передние ножки свода (crura anteriora fornicis) спускаются впереди зрительных бугров, соединяясь с ними, к основанию мозга и здесь соединяются со сосковидными телами; задние (crura posteriora fornicis) — направляются в нижние рога боковых желудочков, к Аммониеву рогу. Берега щели 3-го желудочка впереди, посередине и сзади соединяются между собою тремя полосками мозговой ткани, наз. передней, средней, или серой, задней спайками (commissurae anterior, mediagrisea, posterior). Полость третьего желудочка сообщается с боковыми посредством отверстия (foramen Monroi) под передней спайкой; с Сильвиевым водопроводом — посредством отверстия (aditus ad aquaeductum) под задней спайкой; с воронкой основания мозга — посредством отверстия (aditus ad infundibulum) в передней половине дна желудочка. Все описанные мозговые желудочки, равно как и Сильв. водопр., выстланы одним слоем эпителиальных клеток, снабженных у молодых индивидуумов ресницами. Этот слой клеток в желудочках называется эпендимой. Продолжение этого же слоя клеток образует верхнюю стенку желудочков. Во всех описанных желудочках под этим слоем лежит отросток мягкой мозговой оболочки (см. ниже) — только в передний и задний рог боковых желудочков мозговая оболочка не заходит.
Между передними ножками свода и головкой мозолистого тела натянуты две вертикально стоящие пластинки, наз. прозрачными перегородками (septum pellucidum); они срастаются между собою, но не по всей поверхности; между ними остается полость, наз. Дункановой полостью, или желудочком прозрачной перегородки. Кзади от 3-го желудочка, над переднею частью Варол. моста и ножками мозга, лежит четверохолмие (corpus quadrigeminum); оно состоит из белого вещества, двумя взаимно перпендикулярными бороздами делящегося на четыре холмика, от которых отходят валики (brachia). Валики двух передних холмиков направляются к зрительным буграм и соединяются с передним коленчатым телом; задние валики постепенно сглаживаются по направлению к нижележащим ножкам мозга. Бугорок между передними и задними валиками наз. задним (внутренним) коленчатым телом (corpus geniculatum posterius). Каждый холмик четверохолмия и заднего коленчатого тела заключает в себе по серому ядру. На четверохолмии между его передними бугорками лежит буроватое тельце конической формы, вершиной обращенное кзади; оно наз. шишковидной железой, или надмозговым придатком (glandula pinealis, epiphysis), и состоит из пузырьков, наполненных клеточками. Мы уже говорили, что серое вещество образует поверхностный слой полушарий большого мозга, называемый коркой; толщина последней равна 2-5 мм. Лежащее под коркой белое вещество, наз. полуовальным центром (centrum semiovale Vieussenii), заключает в себе следующие большие серые узлы; кнаружи от хвостатого тела (nucleus caudatus) лежит в массе белого вещества полушария большой узел двояковыпуклой формы, наз. чечевичным ядром (nucleus lenticularis). Слой белого вещества, отделяющий чечевицу от хвостатого тела, наз. внутренней капсулой (capsula interna); внутренняя капсула изогнута под углом, открытым кнаружи, и есть не что иное, как продолжение ножек мозга, вступивших в последний. Кнаружи от чечевицы лежит наружная сумка (capsula externa), кнаружи от последней — вертикальная пластинка серого вещества, наз. оградой (claustrum). Эти три ядра (nucleus caudatus, n. lenticularis, n. claustrum) образуют полосатое тело (corpus striatum). В височной доле полушария — серое ядро величиною с горошину, наз. миндалиной (nucleus amygdalae, amygdala). Зрительные бугры и полосатые тела сливаются своими нижними поверхностями с серым веществом основания мозга, почему и могут быть рассматриваемы как разращения корки, направляющиеся в полуовальный центр. Г. мозг дает начало 12 выходящим из правой и левой его половины черепным нервам. Эти нервы начинаются корешками, т. е. пучками нервных волокон от тех или иных узлов серого вещества, наз. в этих случаях ядрами, центрами нервов; ядра нервов находятся, в свою очередь, в связи с определенными участками коры большого мозга, служащими первичными, главными психическими центрами головных нервов. Психические центры находятся в перекрестном отношении с телом, т. е. какая-нибудь часть, орган правой половины тела связана посредством нервных волокон с левыми психическими центрами. По-видимому, только обонятельные нервы составляют исключение из общего правила: правый центр с правою половиной носовой полости, левый — с левой. Все 12 пар нервов отходят от нижней поверхности головного мозга. 1-я пара нервов — обонятельный нерв (n. olfactorius) — начинается тремя белыми полосками от передней продырявленной пластинки, которые соединяются в ствол, лежащий на нижней поверхности лобной доли и, пройдя вперед см на два, утолщающийся в обонятельную луковицу (серого цвета), от которой отходят веточки, направляющиеся через отверстия решетчатой кости в носовую полость. О второй паре — зрительных нервах (nn. optici) — y же было сказано. III пара — нервы, движущие глазом (n. oculomotorii), — отходит от внутреннего края ножек мозга. IV пара — блоковые нервы (nn. trochleares) — начинается между ножкою мозга и височною долей. V пара — тройничные нервы (n. trigemini) — отходит от ножек мозжечка к мосту. Между задним краем Варолиева моста и пирамидой выходит VI пара нервов наружной отводящей мышцы глаза (n. abducens). Между оливой и задним краем Варол. моста — VII пара личных нервов (n. faciales) и VIII пара — слуховые нервы (n. acustici). IX пара — языкоглоточных нервов (n. glosso-pharyngeus) — отходит от боковой поверхности веревчатого тела. Х пара блуждающих, или легочно-желудочных, нервов (n. vagus, pneumo-gastricus) — отходит 10-12 нитями от боковой поверхности продолговатого мозга. XI пара прибавочных нервов (n. accessorius Willisii) — начинается от шейной части спинного мозга, вступает в черепную полость и здесь соединяется с блуждающим нервом, от которого получает ветви. Из бороздки между пирамидой и оливой выходит XII пара подъязычных нервов, иначе движущих нервов языка (n. hypoglossus, motorius linguae). Дальнейшие подробности см. под соответственными названиями нервов.
2) Оболочки мозга. Головной мозг покрыт тремя оболочками (velamenta cerebri), из которых самая внутренняя — мягкая, или сосудистая (pia mater, meninx vasculosa), так как содержит множество сосудов, которые из нее направляются в мозг. Мягкая оболочка входит во все щели и бороздки, а также дает отростки в желудочки. Один такой отросток вступает между мозолистым телом и лежащими под ним четверохолмием и зрительными буграми в третий желудочек и здесь наз. верхним сосудистым сплетением; его продолжение через Монроево отверстие в боковые желудочки дает боковые сплетения. Другой отросток входит в 4-й желудочек и называется нижним сплетением. Над мягкой лежит паутинная мозговая оболочка (arachnoidea mater, meninx serosa); эта не заходит в борозды, щели и прочее, близко лежит на предыдущей и даже срастается с ней на выдающихся частях мозга — на извилинах, буграх и пр. В остальных местах между обеими оболочками остаются узкие, сообщающиеся между собою полости, называемые паутинными пространствами (spatia Subarachnoidalia), заключающие в себе небольшие количества сыворотки (liquor cerebro-spinalis). Наконец, над паутинною оболочкой лежит твердая мозговая оболочка (dura mater, meninx fibrosa), плотно пристающая к шероховатостям, ребрам и пр. внутренней поверхности черепных костей, для которых служит периостом (см.). Твердая оболочка состоит из двух листков, в определенных местах отстающих друг от друга, образуя полости, наз. пазухами (sinus durae matris). От твердой об. отходят несколько отростков, которые делят черепную полость как бы на камеры и защищают отдельные части мозга от давления друг на друга при различных положениях тела. Один из них, наз. большим серповидным отростком (processus falciformis major), представляет вертикально стоящую пластинку, изогнутую и прикрепляющуюся к средней продольной линии черепной крышки; она входит в щель между полушариями большого мозга и передним концом прикрепляется к решетчатой кости, а задним — к мозжечковому намету. Последний, назыв. также палаткой (tentorium cerebelli), есть также отросток твердой оболочки, который вдвигается в щель между полушариями большого и малого мозга. Малый серповидный отросток — узкая пластинка, один конец которой прикрепляется к нижней поверхности палатки, а другой — у задней окружности затылочной дыры. Мозговые оболочки состоят из различных видов соединительной ткани. Все три мозговые оболочки продолжаются и на выходящие из черепной полости нервы.
3) Кровообращение в головном мозгу. Артериальную кровь доставляют правая и левая внутренние сонные артерии (arteria carotis interna) и две позвоночные артерии (arteria vertebralis) — ветви подключичной. Внутренняя сонная делится на переднюю и среднюю мозговые артерии; первая ветвь разветвляется в мозолистом теле и внутренней поверхности полушария; вторая по Сильвиевой борозде разветвляется и на наружной поверхности полушария. Обе артерии идут вдоль продолговатого мозга, дают множество ветвей для мозжечка, червячков и пр., сливаются, образуя основную артерию, лежащую в бороздке Варол. моста; основная артерия кпереди делится на две задние мозговые артерии, которые огибают ножки мозга, поднимаются вверх и снабжают кровью задние доли полушария. Последние две артерии, соединяясь с двумя передними мозговыми артериями, образуют на основании мозга сосудистый кружок (circulus arteriosus Willisii). Из капилляров мозга составляются небольшие венозные стволики, направляющиеся к мягкой мозговой оболочке; здесь они образуют сплетение, находящееся в связи с венозным сплетением спинного мозга. Из мягкой оболочки венозная кровь изливается в пазухи твердой мозговой оболочки, — это каналы с просветом 3-угольной или круглой формы, помещающиеся то в описанных отростках твердой мозговой оболочки, то в других частях последней. Все пазухи изливают свою кровь в пазуху мозжечковой палатки, назыв. поперечным восприемником (sinus transversus, confluens sinuum); отсюда кровь выводится из черепной полости внутренними яремными венами. Посредством соединений пазухи твердой оболочки образуют венозный кружок (sinus circularis Ridleyi), соответствующий описанному артериальному кружку. В пазухи твердой мозговой оболочки изливают свою кровь также вены оболочек мозга, черепных костей, отчасти вены глаза, носа, уха.
4) Ткань мозга. Элементы и серого, и белого вещества заложены в поддерживающей ткани, называемой невроглией; в белом веществе в невроглии залегают почти исключительно мякотные нервные волокна, в сером веществе в невроглии залегают главным образом нервные клетки, затем нервные нити, голые осевые цилиндры и мякотные нервные волокна. Невроглия имеет сетчатое строение; ее клетки с большими круглыми ядрами и незначительным количеством протоплазмы имеют отростки, образующие сеть, окутывающую нервные клетки и волокна. На поверхности мозга и вблизи желудочков невроглия скопляется в значительном количестве. Каждое нервное волокно берет начало от нервной клетки или на пути своем вступает в связь с нервными клетками того или другого скопления серого вещества. Происходит это или таким образом, что отросток нервной клетки, облекаясь мякотью, становится осевым цилиндром мякотного нервного волокна (этот отросток только в самом своем начале дает несколько тончайших отпрысков, посредством которых, вероятно, нервные клетки соединяются друг с другом), — или же отросток нервной клетки, многократно ветвясь, образует тончайшую сеть (Герлаховская сеть), которая переплетается и соединяется с другой сетью, происшедшей от древовидных разветвлений подходящего нервного волокна. В корке большого и малого мозга серое вещество состоит из нескольких отдельных слоев, отличающихся своими клеточными элементами. Последние — или пирамидальной формы (величиною 0,006-0,120 мм), или, что реже, веретенообразной (0,010-0,012 мм), кругловатой и многоугольной (0,008-0,010 мм). Каждая пирамидка имеет несколько отростков, из которых отходящий от основания пирамиды — осецилиндровый (см. выше), остальные протоплазматические; из последних — отходящий от верхушки пирамидки направляется вверх к свободной поверхности мозга. Самый наружный слой корки большого мозга образует невроглия. Под этим слоем — слой маленьких пирамид; 3-й слой — больших пирамидальных клеток. 4-й — слой круглых клеток (зерен), среди которых встречаются и малые пирамиды; 5-й и последний (внутренний) слой корки образуют веретенообразные клетки. Большие (величиною до 0,120 мм) пирамидальные клетки встречаются главным образом в коре обеих центральных извилин и парацентральной дольки — в коре затылочной и височной долей их мало. В некоторых частях коры число слоев доходит до восьми. Мякотные нервные волокна в корке полушарий большого мозга идут в радиальном направлении (к свободной поверхности полушария) до слоя больших пирамид включительно; в последнем нервное волокно, теряя мякотную оболочку, вступает в большую пирамиду. В слое малых пирамид нервные волокна очень тонки и образуют неправильные сплетения; в поверхностном слое — большинство волокон идет параллельно свободной поверхности; их немного. В самом внутреннем слое волокна и веретенообразные клетки параллельны поверхности мозга. В корке мозжечка — наружный слой образует невроглия, в которой залегают мякотные нервные волокна и отростчатые клетки. Второй слой образует ряд больших нервных клеток, снабженных двумя отростками, из которых один, направляющийся в нижележащий слой, — осецилиндровый; другой, толстый, разветвляясь, направляется к свободной поверхности мозжечка. Третий и последний слой, зерновой, состоит из клеток — зерен с большим ядром и бедных протоплазмой, веретенообразных клеток с отростками и звездчатых клеток невроглии. Белое вещество Г. мозга, как сказано, состоит из мякотных нервных волокон.
5) Ход волокон в мозге. С целью представить в общих чертах взаимные отношения серого клеточного вещества и нервных волокон, а также и ход последних Мейнертом составлена следующая краткая схема: нервные окончания в органах чувств и нервные окончания в мышцах связаны проводниками — чувствительными и двигательными нервными волокнами с коркой полушарий большого мозга; поэтому последняя представляет сумму всех наших отношений к внешнему миру ("корка — проекционная поверхность внешнего мира"). От этой проекционной поверхности идут проводники (проекционная система волокон) к концевым нервным аппаратам. Проекционная система состоит из трех отрезков, или членов: 1) от клеток коры до клеток больших узлов большого мозга (полосатое тело, чечевица, зрительный бугор, четверохолмие); с клетками этих узлов вступает в соединение первый член проекционной системы, который образуют нервные волокна белого вещества полушарий большого мозга; 2) от клеток этих (субкортикальных) узлов до клеток серого вещества полостей (3-го желудочка Сильвиева водопровода, 4-го желудочка, канала спинного мозга), с которыми опять вступают в соединение. Эта часть проводников, перекрещивающихся на своем пути, образует ствол головного и спинного мозга; 3) от серого вещества полостей до периферических нервных окончаний; он образует все черепные и спинномозговые нервы. Кроме проекционной системы волокон, в головном и спинном мозге существуют волокна ассоциирующие и комиссуральные (см. ниже). Волокна мозжечка благодаря его ножкам присоединяются ко второму отрезку проекционной системы. Распределение волокон не настолько изучено, чтобы можно было расположить все волокна Г. мозга по этой схеме. В стволовой части мозга (к которой причисляют все части Г. мозга, за исключением полушарий большого мозга, называемого плащом, или мантией) различают волокна основания ствола и волокна покрышки последнего. К волокнам основания принадлежат: А) волокна пирамид продолговатого мозга, служащих продолжением прямых и боковых пирамидных пучковых спинного мозга. В Варолиевом мосте к этим волокнам присоединяются: В) волокна двигательных нервов, С) пучок чувствующих нервов из петли покрова (см. ниже) и D) волокна мозжечка, попавшие в Варол. м. через ножки мозжечка и образующие два пучка; из них один (е) от коры верхней поверхности мозжечка к коре затылочной доли, другой (f) от коры нижней поверхности мозжечка к лобной доле большого мозга. Все перечисленные волокна из Варол. м. направляются в ножки мозга, где размещаются следующим образом: А и В ложатся в средней трети основания ножки, f — y внутреннего, а е и С — у наружного его края. Все остальные волокна Г. мозга образуют покрышку ствола и попадают в полушария большого мозга через покрышку ножки; ход их недостаточно изучен. Волокна эти лежат в продолговатом мозге и Варол. м. над волокнами пирамид и под серым веществом 4-го желудочка и Сильвиева водопровода. Большинство из них идет в продольном направлении, не образуя компактных пучков; последних только два — в Варол. м.; один лежит в виде широкой полосы, называемой петлей (lemniscus), над волокнами пирамид, а другой, называемый задним продольным пучком, — тотчас под Сильвиевым водопроводом. К волокнам, имеющим поперечное направление, принадлежат: в Варол. м. — волокна ножек мозжечка к мосту; в продолговатом мозге — волокна дугообразные, которые или начинаются от собственного его серого вещества, или представляют продолжения задних пучков спинного мозга. Поперечные и отчасти продольные волокна на своем пути в продолговатом мозге и Варол. м. перекрещиваются в срединной плоскости последних, образуя так назыв. шов (raphe). Продольные и поперечные пучки нервных волокон покрышки ствола вместе с рассеянным серым веществом образуют так называемое сетчатое образование (formatio reticularis), которое тянется от нижнего конца продолговатого мозга до зрительных бугров. Относительно значения и окончания волокон покрышки ствола известно, 1) что через ножки мозжечка к продолговатому мозгу направляются к коре мозжечка: а) мозжечковые пути боковых столбов спинного мозга, b) часть клиновидных и тонких пучков; из последних часть образует предварительно некоторые дугообразные волокна в продолговатом мозге, а потом уж идут в веревчатые тела; с) волокна из ядра олив; 2) что в петлю вступают волокна: а) из основных пучков переднего и бокового столбов спинного мозга, b) из Голлевских и Бурдаховских его пучков, с) некоторые дугообразные волокна продолговатого мозга; они оканчиваются, вероятно, в зрительных буграх, передних холмиках четверохолмия и коре теменной доли большого мозга; 3) задние продольные пучки Варолиева моста несут волокна из основных пучков передних и боковых столбов спинного мозга, вероятно, к зрительным буграм; 4) ножки мозжечка к четверохолмию перекрещиваются над последним, вступают в связь с так называемым красным ядром ножки мозга, откуда поворачивают к теменной доле. Что касается состава внутренней капсулы полушарий, служащей продолжением ножек мозга, то в переднем ее бедре проходит мозжечковый пучок и пучок от зрительных бугров — оба к лобной доле; в колене капсулы проходят А и В (см. выше); в задней трети заднего бедра — (чувствительные) волокна покрышки ствола (carrefour sensitif Шарко).
В большом мозге различают волокна: 1) лучистой системы (corona radiata), 2) ассоциационные, 3) комиссуральные. К лучистой системе принадлежат волокна, которые от внутренней капсулы в виде множества лучей направляются к коре передних и задних долей полушарий; далее — волокна, отходящие в виде лучей от зрительных бугров к коре лобной, теменной и затылочной долей и к коре скрытой дольки мозга; лучи от чечевицы к коре лобной и теменной долей; лучи от головки хвостатого тела к чечевице и отчасти к коре. Все эти волокна в коре полушарий вступают в соединения с ее клетками. К acco циационным принадлежат волокна от верхушки коры какой-нибудь извилины, спускающиеся вниз и опять поднимающиеся к верхушке соседней извилины и пр. Эти волокна назыв. собственными волокнами большого мозга (fibrae propriae). Более отдаленные части коры сочетаются при помощи пучков волокон — так при помощи дугообразного пучка (fasciculus arcuatus) ассоциируется кора лобной доли с корой затылочной и височной. Комиссуральные волокна соединают одноименные части полушарий; сюда принадлежат волокна, отходящие с боков мозолистого тела и волокна спаек 3-го желудочка. Белое вещество мозжечка образуют волокна трех его ножек, волокна ассоциирующие, комиссуральные, волокна корешков нервов.
6) Вес головного мозга — в среднем равен у мужчин 1360-1375 с колебанием от 1018-1925 гр.; у женщин 1220-1245 гр. с колебаниями от 820-1565. После быстрого роста в течение первого года жизни (мозг новорожденного 410 гр. — 1/8 веса тела; вес мозга в конце первого года 900 г p. = 1/14 веса тела) мозг растет медленно и между 20-30 годами достигает предела своего роста, до 50 лет не изменяется, а затем начинает уменьшаться в весе. Между всеми животными человек обладает наибольшим весом мозга, не только относительным, но и абсолютным. Только у кита при длине тела в 10 саж. и весе до 3000 пд. мозг несколько тяжелее человеческого (2816 гр.). Мозг лошади весит 680 гр.; быка — 500 гр.; льва — 250; антропоморфных обезьян 350-400 гран., редко более. Однако больший или меньший вес мозга у различных людей сам по себе не может служить указанием на размеры их умственных способностей; чрезвычайно тяжелые мозги (в одном случае до 2222 гр.) были находимы у ничем не выдававшихся людей. С другой стороны, люди выдающихся способностей нередко отличаются весом мозга, далеко превосходящим средний. Так, мозг Тургенева весил 2012 гр.; Кювье — 1861 гр.; Байрона — 1807; математика Гаусса — 1492; Скобелева — 1451; химика и композитора Бородина — 1383. В то же время мозг Гамбетты весил всего 1160 гр. Богатство психической организации находится в зависимости от количества и качества нервных клеток коркового слоя полушарий и, вероятно, от количества ассоциационных волокон большого мозга.
Ю. Итин.
В. Эмбриональное pa звитие головного мозга. Зачаток нервной системы появляется у всех позвоночных в виде эктодермического впячивания в форме борозды или желоба, на спинной стороне тела; отделяясь от поверхностного эктодермического эпителия, это впячивание замыкается в трубку, лежащую под эктодермой и над спинной струной. Передний конец этой нервной трубки, расширяясь, дает начало Г. мозгу; первоначальный зачаток Г. мозга состоит из трех пар лежащих друг за другом пузырей (пузыревидных расширений переднего конца мозговой трубки, первичных мозговых пузырей) переднего, среднего и заднего (фиг. 1).
ГОЛОВНОЙ МОЗГ
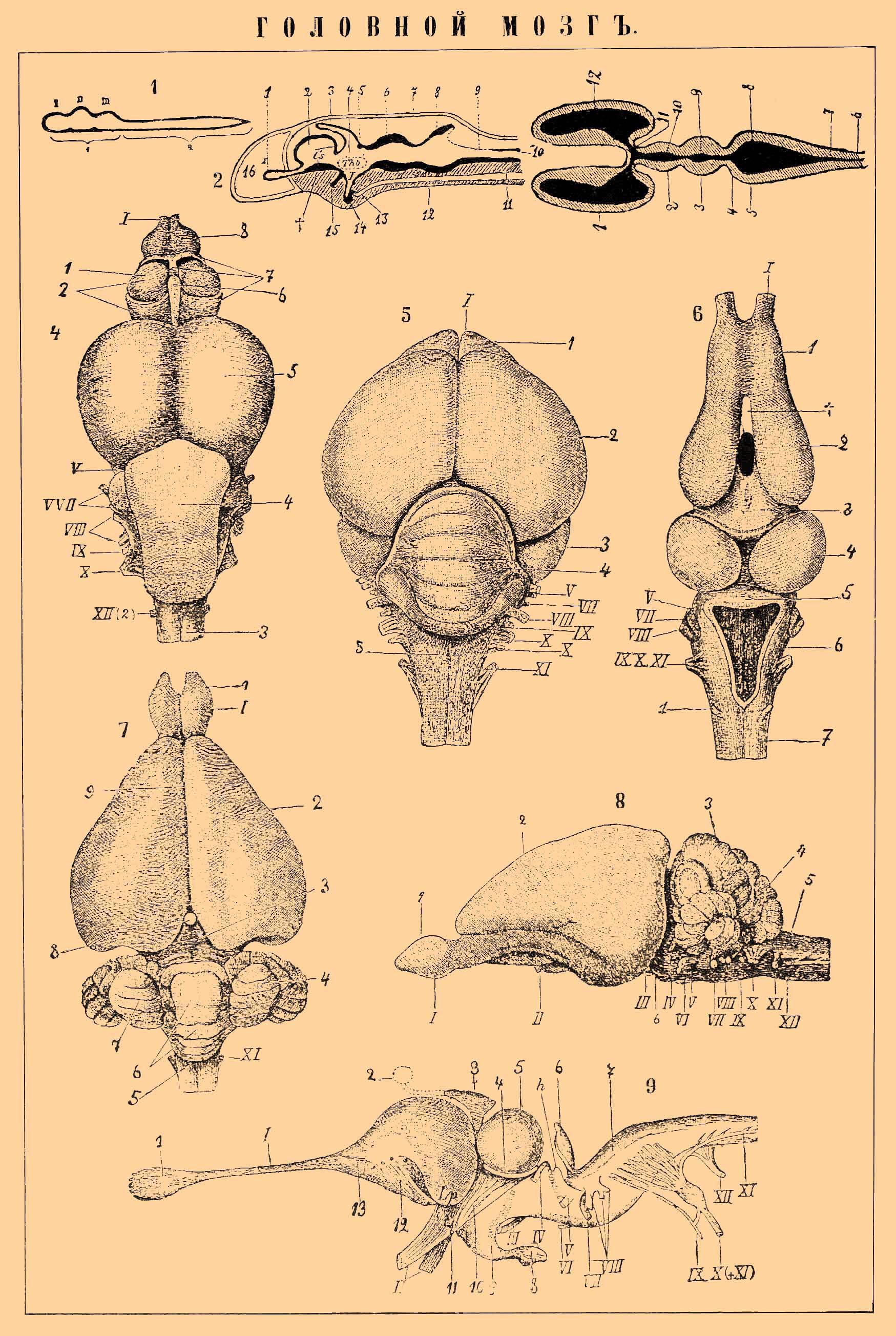
1. Схема эмбрионального развития центральной нервной системы. а — головной мозг с его тремя первичными пузырями (I, II, III); b — спинной мозг.
2. Идеальный продольный разрез через череп и мозг зародыша позвоночных. 1 — обонятельные доли; 2 — передний мозг; 3 — надмозговой придаток; 4 — промежуточный мозг; 5 — задняя комиссура; 6 — средний мозг; 7 — черепная крышка; 8 — задний мозг; 9 — добавочный мозг; 10 — canalis centralis спинного мозга; 11 и 13 — спинная струна; 12 — основание черепа; 14 — подмозговой придаток; J — воронка (infundibulum); Cs — полосатые тела; Tho — зрительные бугры; 15 — зрительный нерв; 16 — носовая полость.
Римские цифры везде обозначают головные нервы (I — обонятельный; II — зрительный; III — n. oculomotorius; IV — n. trochlearis; V — тройничный; VI — n. adbucens; VII - лицевой; VIII — слуховой; IX — glossopharyngeus; X — блуждающий; XI — n. accessorius Willisii; XII — n. hypoglossus.).
3. Схема желудочков головного мозга. 1 — вторичный передний мозг (полушария большого мозга) с (12) боковыми желудочками; 2 — промежуточный мозг с (10) третьим желудочком; 11 - foramen Monroi; 3 — средний мозг и (9) Сильвиев водопровод; 4 — задний мозг; 5 — добавочный (продолговатый) мозг и 8 — четвертый желудочек, который продолжается в (7) центральный канал спинного мозга (6).
4. Мозг костистой рыбы (форели, Salmo fario). 2 — передний мозг; 7 — раllium (мантия) переднего мозга; 1 — основной узел переднего мозга; 6 - glandula pinealis; 5 — средний мозг; 4 — задний (малый) мозг; 3 — добавочный (продолговатый) мозг.
5. Мозг лягушки (Rana esculenta). 1 — обонятельные доли; 2 — передний (большой) мозг; t — свободный промежуток между его полушариями; 3 — промежуточный мозг; 4 — средний мозг; 5 — задний (малый) мозг; 6 — добавочный (продолговатый); 7 — спинной мозг.
6. Мозг ящерицы (Hatteria punctata). 1 — обонятельная луковица; I — обонятельный нерв; 3 — надмозговой придаток, оканчивающийся (2) теменным глазом; 5 — средний мозг; 4 — кольцевая складка при его основании; 6 — малый мозг; 7 — продолговатый; 9 — воронка; 8 — подмозговой придаток; 10 — tractus nervi optici; 11 — chiasma; II — зрительный нерв; 12 — pedunculi cerebri; 13 — передний мозг.
7. Мозг голубя: 1 — обонятельные доли; 2 — большой мозг; 3 — средний мозг; 4 — малый; 5 — продолговатый.
8. Мозг кролика, сверху. 1 — обонят. доли; 2 — большой мозг; 3 — средний мозг (четверохолмие); 5 — продолговатый; 6 — средний отдел малого мозга (червячок, vermis); 4 и 7 — боковые части (полушария) малого мозга; 8 — glandula pinealis; 9 — fissura pallii (щель, разделяющая полушария большого мозга).
9. То же, с боку. Здесь 3 — обозначает малый мозг, 4 — его червячок, 6 — область Варолиева моста. Остальные буквы те же самые.
Римские цифры везде обозначают головные нервы (I — обонятельный; II — зрительный; III — n. oculomotorius; IV — n. trochlearis; V — тройничный; VI — n. adbucens; VII - лицевой; VIII — слуховой; IX — glossopharyngeus; X — блуждающий; XI — n. accessorius Willisii; XII — n. hypoglossus.).
Заключающаяся в них полость соответствует будущим полостям желудочков мозга и сообщается с остальной полостью нервной трубки, следы которой сохраняются в виде центрального канала спинного мозга. Передний и задний первичные пузыри распадаются, в свою очередь, на два отдела каждый, и так происходят 5 основных отделов Г. мозга: 1) передний мозг; 2) промежуточный; 3) средний; 4) задний, и 5) добавочный (Prosencephalon, Thalamencephalon, Mesencephalon, Metencephalon, Муеlеnсерhаlоn) — ф. 2 и 3. Передний мозг, обыкновенно с продольной складкой, дает начало полушариям большого мозга с боковыми желудочками; в основании его образуется ганглиозная масса (полосатые тела, corpora striata), составляющая ядро большого мозга; остальная часть его стенок образует мантию, или плащ (раllium). От переднего мозга отходят обонятельные лопасти (lobi olfactorii). Промежуточный мозг утолщением стенок дает начало зрительным буграм (thalami optici), между которыми сохраняется первоначальная полость пузыря в виде третьего желудочка; последний сообщается с обоими боковыми желудочками через Монроевы отверстия (foramen Monroi). В виде выпячивания стенок промежуточного мозга образуются первичные глазные пузыри, из которых происходит сетчатка глаза и ее пигментный эпителий; на месте стебельков, соединяющих глазные пузыри с промежуточным мозгом, на передней границе последнего, образуются зрительные нервы, делающие при самом начале перекрест (chiasma). Позади перекреста дно третьего желудка выпячивается в конический вырост, воронку (infundibulum), слепой конец которой образует заднюю лопасть подмозгового придатка (hypophysis cerebri); передняя лопасть последнего образуется на счет эпителия первичного ротового углубления. Верхняя стенка промежуточного мозга, составляющая крышку третьего желудочка, сохраняет характер тонкой перепонки; на ней развивается особый вырост — надмозговой придаток, или шишковидная железа (ерi physis cerebri s. glandula pinealis). Оба передние отдела мозга лежат впереди хорды. Средний мозг делится на два или четыре бугра (corpora bigemina, С. quadrigemina — четырехолмие). Задний мозг, который только на спинной стороне явственно отделяется от добавочного, образует малый мозг, или мозжечок (cerebellum), а добавочный мозг превращается в продолговатый (medulla oblongata), непосредственно продолжающийся в спинной. Общая полость заднего и добавочного мозговых пузырей составляет четвертый желудочек, назади переходящий в центральный канал спинного мозга, а спереди, посредством Сильвиева водопровода — внутренней полости среднего мозга — сообщается с третьим желудочком. Покров 4-го желудочка, образуемый в передней части мозжечком, в задней части (в продолговатом мозге) состоит из тонкого слоя эпителия, без нервного вещества (как и крышка третьего желудочка). От этого полость 4-го желудочка кажется открытой сверху, образуя ромбоидальную ямку (fossa rhomboidalis). Подмозговой и надмозговой придатки (epiphysis и hypophysis) принадлежат к рудиментарным органам с неизвестной функцией и недостаточно выясненным морфологическим значением (о значении epiphysis как рудимента 3-го глаза см. ниже).
В разных классах позвоночных животных одни из названных 5 отделов Г. мозга разрастаются более других и частью или вполне их прикрывают. Общая масса Г. мозга постепенно возрастает от низших позвоночных к высшим по сравнению со спинным мозгом и массою тела. Особенно важное значение принадлежит полушариям большого мозга, которые постепенно увеличиваются в размерах и наконец у человека покрывают собою все прочие части головного мозга. По мере развития большого мозга, а особенно его коркового слоя совершенствуются и психические способности животных. Корковый слой в мозговой мантии (pallium) существует не у всех позвоночных, и там, где он уже образовался, он не всегда покрывает весь мозг. У миног (Cyclostomata), у костистых рыб (Teleostei) и ганоидных рыб pallium состоит из простого эпителиального слоя; но и у тех позвоночных, где он состоит из нервной ткани, его задний край, прилегающий к промежуточному мозгу, сохраняет эпителиальный характер и впячивается мягкою мозговою оболочкою (pia mater) в полость переднего мозга, образуя plexus chorioideus. Настоящий, трехслойный, с пирамидальными клетками корковый слой больших полушарий появляется лишь у пресмыкающихся, и у всех высших позвоночных с его дальнейшим развитием связано развитие психических способностей. У млекопитающих увеличение количества серого вещества достигается путем увеличения поверхности большого мозга посредством извилистых складок, образующих извилины (gyri) с разделяющими их бороздами (sulci); более тесное соединение обоих полушарий достигается у млекопитающих образованием обширных комиссур в виде мозолистого тела (corpus callosum) и свода (fornix).
С. Строение головного мозга в различных классах позвоночных животных. У простейшего из позвоночных, у ланцетника (Аm phioxus), спинной мозг на переднем своем конце, за которым хорда еще продолжается вперед, не представляет расширений, которые можно было бы приравнять к Г. мозгу. Г. мозг рыб в большинстве случаев занимает лишь часть черепной полости, остальное пространство которой выполнено жиром и лимфой. У селахий (акуловые рыбы) передний мозг отличается относительно значительной величиной и более или менее ясно разделяется на два полушария: боковые желудочки отделены друг от друга вертикальной перегородкой и продолжаются спереди в обонятельные лопасти (lobus olfactorius), соединенные с передним мозгом посредством более или менее длинных стебельков (tractus olfactorius). Мозговой плащ состоит из нервного вещества. Промежуточный мозг акул имеет вид узкой комиссуры между передним и средним мозгом. Спинная стенка его, составляющая крышку третьего желудочка, вытягивается наперед в трубкообразный epiphysis, нередко заходящий далее переднего конца мозга, и передним концом вдается в черепную крышку. На брюшной стороне промежуточного мозга, по бокам воронки и hypophysis, лежат две нижние лопасти (lobi inferiores), в которых продолжается полость воронки, а позади их лежит пара сосудистых мешков (sacci vasculosi), полость которых также сообщается с воронкой. Мозжечок имеет значительный объем, так что отчасти прикрывает отделы, лежащие впереди и сзади его. Объемистый продолговатый мозг отличается тем, что боковые стенки ромбоидальной ямки представляют вздутия, вдающиеся в полость 4-го желудочка; из них передние соответствуют началу тройничного нерва и называются его лопастями (lobi trigemini), a задние соответствуют началу блуждающего нерва (lobi nervi vagi). У гнюса, или электрического ската (Torpedo), существуют еще особые электрические лопасти (lobi electrici), выполняющие всю ромбоидальную ямку и заключающие в себе гигантские ганглиозные клетки. Г. мозг костистых рыб (Teleostei) (фиг. 4) замечателен тем, что передний мозг представляет непарный пузырь, раllium (мозговой плащ) которого лишен нервных элементов и состоит лишь из простого слоя эпителиальных клеточек, тогда как нижняя, брюшная стенка образует два значительные утолщения, вдающиеся в общую полость (ventriculus communis) переднего мозга и соответствующих частям полосатых тел (с. striata). Тонкая оболочка мозга (рiа mater) впячивает крышку переднего мозга спереди назад, в общий желудочек, в виде поперечной складки, образуя сосудистое сплетение. Стенки epiphysis имеют такое же строение, как и крышка переднего мозга. Промежуточный мозг обыкновенно не заметен сверху, нижние лопасти его представляют более значительное развитие, чем у селахий. Позади hypophysis лежит сосудистый мешок ( saccus vasculosus), представляющий из себя разветвленную железу, выводной проток которой открывается в воронку. Средний мозг разделен сверху на два полушария (lobi optici). Задний мозг своею верхнею стенкою более или менее впячивается в полость среднего мозга. Вообще Г. мозг костистых рыб значительно уступает по размерам Г. мозгу селахий, и самым крупным отделом его являются средний и задний мозг. — Между ганоидными рыбами по строению мозга одни приближаются к костистым рыбам, другие к двоякодышащим (Dipnoi). Г. мозг последних парными, хорошо развитыми полушариями большого мозга, отсутствием нижних лопастей (lobi inferiores) и слабым развитием заднего мозга (мозжечка), имеющего вид узкого поперечного мостика на переднем краю ромбоидальной ямки — напоминает амфибий. Г. мозг амфибий, особенно бесхвостых, вообще напоминает еще рыб незначительною величиною и расположением своих отдельных частей почти в горизонтальной плоскости (фиг. 5, 6). Полушария большого мозга удлиненные и напереди непосредственно переходят в обонятельные лопасти. У хвостатых амфибий оба полушария по всей длине отделены друг от друга и соединены лишь узкой комиссурой впереди Монроева отверстия; у бесхвостых они разделены в задней части и опять сливаются между собой обонятельными лопастями. Средний мозг у бесхвостых достигает значительных размеров, разделен сверху на две половины, и его внутренняя полость (aquaeductus Sylvii) дает у лягушек два боковых выступа. Мозжечок очень слабо развит и является в виде узкой поперечной пластинки над ромбоидальной ямкой. Epiphysis y бесхвостых первоначально имеет вид полого отростка, конец которого проходит через черепные кости и оканчивается расширением в виде пузырька под эпидермой кожи; во взрослом состоянии стебелек эпифиза теряет полость и перерождается, но сохраняется на всю жизнь. Мозг гимнофионов отличается высоким развитием, именно значительной величиной передних полушарий (см. Безногие гады).
У пресмыкающихся передний мозг покрывает уже промежуточный и простирается до среднего мозга (фиг. 9). Его полушария обладают довольно большими боковыми желудочками с хорошо развитыми основными узлами и соединены между собой лишь узкой комиссурой (commissura anterior), расположенной впереди третьего желудочка. От крышки третьего желудочка отходит вверх и наперед длинный epiphysis, который у ящериц через теменное отверстие (foramen parietale) выходит из черепной полости и оканчивается в коже головы, прикрытый нередко особыми прозрачными чешуйками. Средний мозг продольной бороздкой разделен на два полушария. Мозжечок у змей и ящериц, как у амфибий, лежит в виде узкой вертикальной пластинки над передним краем ромбоидальной ямки; у черепах и крокодилов он гораздо шире, и особенно у крокодилов его средняя часть отличается величиною и выпуклостью. В строении epiphysis можно различить у пресмыкающихся три типа. У одних он имеет вид длинной трубки, которая сообщается с полостью промежуточного мозга или 3-го желудочка, и оканчивается под кожей головы, над foramen parietale, маленьким пузырьком, выстланным внутри мерцательным эпителием. У других концевой пузырек не имеет сообщения с третьим желудочком и соединен c верхней стенкой промежуточного мозга посредством плотного стебелька, образованного из волокон и веретеновидных клеток и представляющего сходство со зрительным нервом зародышей. Третий тип (у ящериц из родов Наttеriа, Monitor, Lacerta, Anguis) представляет из себя так наз. третий глаз позвоночных. Здесь передняя стенка концевого пузырька epiphysis, лежащая всего ближе к поверхности тела, представляет в середине прозрачное утолщение, вдающееся в полость пузыря наподобие хрусталика (см. рис. при ст. Глаз). Задняя стенка пузыря по строению напоминает ретину; в ней заключаются пигмент и клеточные образования, похожие на палочки сетчатки. Этот орган, вероятно, представляет из себя рудимент особого, непарного, теменного глаза, существовавшего у древнейших позвоночных, но сохранившегося в узнаваемой (и даже, может быть, функционирующей) форме лишь у немногих ящериц, тогда как у остальных позвоночных следы его существуют в различных ступенях атрофии. У некоторых ящериц (Anguis) не один теменной глаз, а 2-3, сидящих на общем стебельке.
У птиц Г. мозг относительно гораздо больше, чем у пресмыкающихся (фиг. 7). Полушария большого мозга соединены между собой передней комиссурой (commissura anterior), залегающей в передней стенке третьего желудочка; сзади нее находится поперечный пучок волокон, расходящихся в оба полушария, и представляющий из себя зачаток мозолистого тела (corpus callosum). Боковые желудочки продолжаются в небольшие обонятельные лопасти. Главную массу большого мозга составляют сильно развитые соr pora striata. Полушария большого мозга простираются назад до мозжечка; зрительные лопасти (lobi optici) среднего мозга сдвинуты в сторону и вниз; у многих птиц они вовсе не заметны сверху и лежат в виде выпуклых бугров на боковых поверхностях и на основании мозга. Epiphysis, лежащий в треугольном промежутке между полушариями большого мозга и мозжечком, имеет вид маленького овального тела, соединенного тонким стебельком с крышкой промежуточного мозга и прилегающего концом своим к твердой мозговой оболочке; он состоит из многочисленных эпителиальных пузырьков, связанных посредством соединительной ткани, богатой кровеносными сосудами. Мозжечок состоит из большей средней части и двух маленьких боковых придатков (flocculi); ромбоидальную ямку 4-го желудочка мозжечок прикрывает вполне, так что продолговатый мозг сверху мало заметен. Средняя часть мозжечка поперечными бороздками разделяется на многочисленные листочки.
У млекопитающих преобладающего развития достигают большой мозг и мозжечок (фиг. 8). У низших млекопитающих (однопроходных, сумчатых, насекомоядных, неполнозубых, грызунов и рукокрылых) полушария большого мозга еще не закрывают сверху среднего мозга (четырехолмий); у хищных и копытных они тянутся назад до мозжечка; у обезьян и человека большая часть последнего покрыта полушариями большого мозга. Поверхность мозговых полушарий у утконоса, у многих сумчатых и неполнозубых такая же гладкая, как у других позвоночных. У остальных млекопитающих на поверхности полушарий появляются извилины и борозды, назначение коих — увеличение поверхности больших полушарий и составляющего ее ганглиозного коркового слоя; относительное развитие ее находится в связи с размерами животного (значительное число извилин у китообразных, копытных и слонов) и с развитием интеллектуальн. способностей. Отодвинутые вниз обонятельные лопасти своим передним вздутым концом обыкновенно несколько выдаются впереди большого мозга; у обезьян, человека и некоторых водных млекопитающих с рудиментарными обонятельными органами (китообразные, тюлени) они прикрыты лобными лопастями полушарий. Боковые желудочки представляют два отдела: обширный передний рог (cornu anterius) с полосатыми телами (corpora striata) и нижний (cornu inferius) с вдающейся в него складкой внутренней стенки полушарий, образующей Аммонов рог (cornu Ammonis, pes hippocampi major); y высших обезьян и у человека существует еще задний рог (cornu posterius) с такою же складкой (pes hippocampi minor, calcar avis). Полушария большого мозга соединены двойною системою комиссур: сводом (fornix) и мозолистым телом, что особенно отличает передний мозг млекопитающих от других позвоночных. Вследствие срастанья обоих полушарий боковые желудочки разделены тонкой перегородкой (septum pellucidum), заключающей в себе щелевидную полость — 5-й желудочек (ventriculus quintus, v. septi pellucidi); тогда как остальные желудочки представляют остаток первоначальной полости мозговых пузырей, 5-й желудочек составляет замкнутую часть первоначальной наружной поверхности мозга. Мозолистое тело у различных млекопитающих развито неодинаково. У однопроходных и сумчатых — это тонкая и короткая пластинка; у грызунов, насекомоядных и др. оно развито сильнее и достигает наибольшего развития у обезьян и человека. Передняя комиссура (commissura anterior), у низших позвоночных составляющая единственное соединение обоих полушарий, сильно развита еще у однопроходных и сумчатых, но у высших млекопитающих имеет вид тонкого шнурка, расположенного впереди свода. Thalami optici (зрительные бугры) промежуточного мозга своими боковыми поверхностями срастаются с corpora striata, так что кажутся принадлежащими к переднему мозгу. Третий желудочек, имеющий вид узкого пространства между зрительными буграми, пересекается посередине средней комиссурой (с. mollis), соединяющей зрительные бугры. Epiphysis у млекопитающих, как и у птиц, никогда не достигает значительного развития, и состоит из многочисленных пузырьков, связанных сосудистой соединительной тканью и выполненных клетками, похожими на лимфатические. Средний мозг отличается небольшими размерами по сравнению с этим отделом мозга у других млекопитающих. Его полость имеет вид узкого канала (Сильвиев водопровод), соединяющего третий желудочек с четвертым. Две бороздки, продольная и поперечная, разделяют поверхность среднего мозга на 4 бугра, образуя четырехолмие (corpora quadrigemina). У однопроходных это разделение выражено весьма слабо. Мозжечoк млекопитающих состоит из среднeго отдела, червячка, (vermis) нередко загнутого вбок, и из боковых лопастей. У однопроходных, подобно тому как у птиц, средний отдел сильно преобладает над боковыми, которые являются в виде незначительных придатков. У сумчатых, неполнозубых, рукокрылых и грызунов средний отдел не уступает боковым. Только у хищных и копытных боковые части становятся больше среднего отдела, образуя полушария мозжечка, а у Primates средний отдел в сравнении с полушариями является уже весьма неразвитым. Поверхность мозжечка представляет поперечные складки, расположенные отдельными группами, образующими многочисленные маленькие лопасти. Подобно полушариям большого мозга, мозжечок состоит из центрального белого вещества и серой коры; на продольном вертикальном разрезе мозжечка белое вещество представляет древовидно разветвленную фигуру (древо жизни, arbor vitae). У человека, где мозжечок особенно богат извилинами, древо жизни отличается многочисленными разветвлениями. В коре мозжечка замечательны расположенные в один ряд крупные, грушевидные клетки (клетки Пуркинье), из которых каждая на внутреннем конце дает тонкий отросток, образующий сперва многочисленные веточки и продолжающийся затем в виде осевого цилиндра в одно из нервных волокон белого вещества, а на противоположном конце посылает несколько сильно ветвистых отростков, направляющихся к поверхности мозжечка. С развитием полушарий мозжечка появляется на нижней поверхности первоначального заднего мозга Варолиев мост в виде широкой поперечной комиссуры, соединяющей оба полушария. У однопроходных и сумчатых в связи с малым развитием полушарий мозжечка слабо выражен и Варолиев мост; у высших обезьян и человека он достигает наибольшего развития. Ср. О головном мозге человека, см. трактаты по анатомии человека: Schwalbe, "Lehrbuch der Neurologie"; Gegenbaur, "Traité d'anatomie humaine, traduit par Julin" (П. 1889); Зернов, "Руководство описательной анатомии человека" (М. 1890); Панш, "Основы анатомии человека". Перев. Таренецкого (СПб. 1888). По эмбриональному развитию мозга см. Hertwig, "Lehrbuch der Entwickelugsgeschichte des Menschen u. der Wirbelthiere" (Иена, 1888; есть русский перевод). По сравнительной анатомии мозга позвоночных см. Бобрецкий, "Основания зоологии" (т. II, 1891); Wiedersheim, "Grundriss der vergleichenden Anatomie der Wirbelthiere" (Иена, 1888; есть русский перевод); его же "Lehrbuch der vergleichenden Anatomie" (Иена, 1886). В большинстве названных сочинений приведена обширная специальная литература.
В. Фаусек.
D. Функции головного мозга крайне разнообразны сообразно с различными отделами его (см. выше анатомию Г. мозга).
1) Продолговатый мозг по функциям приближается более к спинному мозгу, так как не заключает в себе центров произвольной, автоматической деятельности для скелетных мышц всего тела; животное с удаленными вплоть до верхней границы продолговатого мозга всеми отделами Г. мозга по успокоении от раздражения, внесенного операцией, продолжает сидеть все время покойно и отвечает движением только на раздражения. Продолговатый мозг может, след., рассматриваться преимущественно как центр рефлекторных актов и как проводник нервных возбуждений. В первом отношении продолговатый м. является местом локализации многочисленных нервных центров, заправляющих разнообразнейшими функциями тела. Тут лежат центры жевания, сосания, глотания, рвоты, закрывания век; центры дыхательных движений — вдыханий и выдыханий, задерживающие центры сердца, сосудодвигательные центры, центры слюноотделения, желудочного сока и поджелудочного, потоотделения, наконец, центры сложных рефлекторных движений, участвующих в акте передвижения животных, и судорожный центр, вызывающий при своем возбуждении различными ядами или при задушении животных общие конвульсии в теле. Животное, у которого вырезаны все части головного мозга, за исключением только продолговатого, продолжает дышать, способно проглатывать пищу, введенную в глотку, сердце его еще умеряется задерживающим действием блуждающих нервов, кровеносные трубки находятся еще в состоянии тонического сокращения, пищеварительные соки еще могут отделяться рефлекторно и т. д., и стоит только уничтожить продолговатый мозг, чтобы все эти явления исчезли и наступила быстрая смерть. В продолговатом мозгу, именно на дне 4-го желудочка, локализируется Флурансов жизненный узел, разрушением которого прекращается дыхание, вызывается временная остановка сердца и, след., наиболее быстрая смерть животного. Обыкновенно и пользуются уколом в это место для быстрого прекращения жизни. Такое огромное физиологическое значение продолговатого мозга обусловливается еще и тем, что из 12 пар черепных нервов 9 начинаются из него, а именно из дна 4-го желудочка. К продолговатому мозгу прямо направляется целая масса возбуждений, идущих по тройничному, слуховому, языкоглоточному нервам, и в то же время им выводятся непосредственно центробежные возбуждения по двигательным нервам глаз (за исключением только nervus oculomotorius), по двигательным ветвям тройничного нерва, по личному нерву, языкоглоточному, блуждающему, подъязычному и придаточному нервам. Большинство нервных актов, имеющих непосредственным источником своим продолговатый мозг, носит характер чисто рефлекторный, т. е. они вызываются раздражениями, доносимыми до центров из сферы органов чувств и верхних или передних частей Г. мозга; хотя центры дыхательных движений, сердечно-задерживающие центры и сосудодвигательные не делают исключения из этого общего правила подчинения их внешним возбуждениям; тем не менее, они отличаются еще тем общим свойством, что возбуждаются и центрально протекающей по продолговатому мозгу кровью, чем и обусловливается автоматический характер деятельности дыхательных сосудосуживающих и сердечно-задерживающих центров, продолжающих свою работу и помимо всяких внешних возбуждений и волевых импульсов. Продолговатый мозг, связывая спинной мозг с Г., имеет важное значение в качестве проводника нервных возбуждений, с одной стороны чувствующих, направляющихся к Г. мозгу, с другой — центробежных вообще и двигательных в частности, идущих от него к периферии. В нем нервные пути, как двигательные, так и чувствующие, из левой половины спинного мозга переходят в правую половину Г. мозга, а из правой половины спинного — в левую половину Г. мозга. По мнению одних, это перекрещивание обоего рода путей является полным и в таком случае всей левой половиной тела должна была бы заведовать правая половина Г. мозга — правое большое полушарие мозга, а правой — левая половина Г. мозга. Однако доказано, 1) что продольный разрез через место перекрещивания пирамид не вызывает потери волевых движений в обеих сторонах тела, 2) половинная перерезка продолговатого мозга не уничтожает вполне чувствительности в противоположной половине тела, а только ослабляет ее, и то же самое наблюдается и с движениями в этой половине тела, которые частично ослабевают, и, наконец, в-3) частичная потеря произвольных движений, являющаяся вслед за односторонней перерезкой продолговатого мозга, отражается, хотя слабее, и на соответственной половине тела. Очевидно, что перекрест путей представляется неполным. Тут следует припомнить, что часть чувствующих нервных волокон кожи уже перекрещивается в спинном мозгу. Из представленного очерка продолговатого мозга очевидна огромная важность его в явлениях жизни.
2) Мозговые ножки (crura cerebri) и Варолиев мост (Pons Varolii) ввиду обилия в них, кроме белого, серого мозгового вещества, содержащего главным образом нервные центры, должны выполнять какие-нибудь важные нервные функции; к сожалению, известно только то, что мозговые ножки служат главным путем соединения между спинным и Г. мозгом и что оба органа имеют близкую связь со способностью животных производить сложные сочетанные движения вроде ходьбы, бегания, верчения и т. д., так как насильственные движения этого рода происходят нередко при перерезках, повреждениях мозговой ножки и Варол. м. и притом способность эта не утрачивается по удалении полушарий Г. мозга, полосатых тел, зрительных бугров и даже мозжечка. Полагают поэтому, что в Варол. м. в особенности лежат центры локомоторных движений; но тут же должны быть и чувствующие рефлекторные центры, так как животные, лишенные всех частей Г. мозга, за исключением мозговых ножек и Варол. м., отвечают бессознательными криками на щипание кожи. Половинная перерезка Варол. м. часто производит паралич противоположной стороны тела, следовательно, двигательные проводники здесь продолжают еще перекрещиваться; и в то же время может существовать паралич лица, односторонний с поражением Варол. м. и, след., противоположный параличу членов, — факт, объяснимый повреждением лицевого нерва под местом его перекрещивания в Варол. м. О ходе чувствующих путей по Варол. мосту точного известно мало. Мозговые ножки в своей нижней, или брюшной, части (crusta) проводят двигательные пути и стоят в связи с полосатым телом, чечевичным ядром (nucleus lentiformis) и внутренней капсулой; вследствие этого полосатое тело и чечевичное ядро называются ганглиями ножки. Повреждения этой части мозговой ножки вызывают двигательные параличи в накрест лежащей стороне тела. Верхняя, или дорсальная, часть мозговой ножки (tegmentum) проводит чувствующие пути и стоит в связи со зрительными буграми, четырехолмием и внутренней капсулой; поэтому повреждения этой части мозговой ножки должны нарушать чувствительность и вызывать различные формы анестезии. Односторонняя полная перерезка мозговой ножки вызывает после криков боли и конвульсий в накрест лежащей половине тела анестезию и полный паралич этой стороны, тогда как поранения мозговых ножек без перерезки их вызывают ряд насильственных манежных движений. Эти движения, вызываемые также повреждением Варол. м. четырехолмия и мозжечка, наблюдаются не только у животных, но и у человека. Эти различные формы насильственных движений сводятся на: 1) вращение животного вокруг продольной оси тела — при перерезке одной мозговой ножки, одной половины Варол. м., а также продолговатого мозга и четырехолмия; при этом животное вращается то в сторону повреждения, то наоборот. Наблюдалось и у человека. 2) Манежные движения по кругу — при неполных перерезках мозговой ножки или поперечной перерезке Варолиева моста. 3) Движения в форме вращения спиц колеса или часовых стрелок — при неподвижности задней части тела, остающейся как бы в центре, передняя часть круговым образом перемещается — при поранениях передних двухолмий и реже зрительных чертогов и Варол. м. 4) Вращение вокруг поперечной оси тела — кувыркание через голову или прямолинейное неудержимое движение вперед или назад при повреждениях мозжечка и полосатых тел. Все, что можно сказать общего об этих своеобразных движениях, сводится к следующему: побудителями к ним служат, с одной стороны, сильные раздражения и возбуждения, вносимые в центральную нервную систему поранениями ее и поддерживающиеся в течение более или менее долгого времени; а с другой — нарушение проведения к мозгу множества чувствующих возбуждений из сферы кожи, мышц и других органов чувств, столь способствующих поддерживанию равновесия тела и регулирующих всеми движениями его; отсутствие этих центростремительных возбуждений или нарушение правильного подвода их к мозгу должно влечь за собою потерю способности ориентироваться в пространстве и управлять отдельными движениями, и животное, лишенное этих нормальных чувствующих моментов, делается как бы жертвой иллюзий и стремится как бы все одними и теми же движениями восстановить теряемое им все время равновесие.
3) Мозжечок. По опытам Флуранса и др., удаление или повреждение небольших частей мозжечка ведет к неправильностям в походке, в полете, а полное удаление мозжечка совершенно уничтожает координации движений. Односторонняя перерезка средней мозжечковой ножки вызывает насильственные движения, сопровождаемые судорожным вращательным движением глаз. Координирующие двигательные функции мозжечка бывают преимущественно связаны с чувствующими импульсами, подводимыми к нему из полукружных каналов. Несмотря на то, что и у человека наблюдаются при легких поражениях мозжечка неправильности в походке и т. д., тем не менее, известны необъяснимые случаи обширного поражения мозжечка до полного разрушения и не отражавшиеся нисколько на способности людей к сложным локомоторным движениям. Повреждения или удаления боковой доли мозжечка ведут к тем же насильственным движениям, что и перерезка средней мозжечковой ножки (Гитциг и Феррье), а Флуранс заметил, что повреждения средней части мозжечка впереди или назади обусловливает падение животного то вперед, то назад. По опытам Нотнагеля, расстройства координации движений получаются при нарушении непрерывности между обеими половинами мозжечка, тогда как уничтожение целой половины мозжечка у кроликов не дает этих расстройств. Очевидно, что разноречие в показаниях авторов зависит и от индивидуальных особенностей в организации нервной системы у различных видов животных. Феррье прямым возбуждением определенных мест поверхности мозжечка получал у собак и кошек горизонтальное, вертикальное или косвенное движение глаз, иногда с вращением глазных яблок, сопровождаемое движениями головы, конечностей и зрачков. Движения эти вызываются, по-видимому, местным раздражением двигательных центров мозжечка, так как даже механическое раздражение его дает те же результаты (Нотнагель), как и электрическое, и в пользу этого говорит еще и прямая анатомическая связь определенных частей боковых столбов спинного мозга с мозжечком (Флексиг). В связи с описанными выше функциями мозжечка стоят явления, замеченные Пуркинье: при пропускании постоянного тока через голову по направлению от одного уха к другому появляется головокружение, предметы как бы вращаются по направлению тока, и субъекты, подвергаемые электролизации, невольно наклоняются в сторону анода. Гитциг заметил еще, что при известной силе тока появляются, кроме того, колебательные в горизонтальной плоскости движения глаз (nystagmus). Вероятно, ток расстраивает мозжечковый координирующий механизм, из которого посылаются компенсирующие движения глаз и тела, а в сознании это расстройство выражается чувством головокружения. Предполагавшаяся раньше связь мозжечка с половыми функциями (Галль) не оправдалась, и центры эрекции находятся в поясничной части спинного мозга (Гольц); на них-то, вероятно, и действуют возбуждения, исходящие из сферы полушарий мозга; пути же, проводящие эти возбуждения, проходят из полушарий мозга через мозговые ножки и продолговатый мозг.
Наконец, известно, что повреждение известных частей мозга может, подобно повреждению продолговатого мозга, вызывать сахарное мочеизнурение (Экгардт) и что возбуждения мозжечка отражаются и на движениях пищевода и желудка, а повреждения ножек мозжечка нередко ведут к воспалению кишечного канала с кровотечением в нем (Шифф). До сих пор нет удовлетворительной теории, объясняющей механизм координирующего действия мозжечка в локомоторных актах животных.
4) Четырехолмия заключают прежде всего зрительные центры, так как одностороннее разрушение четырехолмия ведет к слепоте глаза или противоположной стороны (у животных с полным перекрестом зрительных нервов), или той же стороны (у животных с неполным перекрестом их). Двустороннее же разрушение четырехолмия ведет к полной слепоте обоих глаз и к постоянному расширению зрачков (Флуранс). Животные с разрушенными большими полушариями Г. мозга при целости четырехолмия продолжают все же быть впечатлительными к свету и движением глаз и головой следят за двигающейся перед ними зажженной свечкой. Только получаемые ими при этом световые впечатления представляются грубыми, несовершенными, лишенными психической окраски. Большая часть волокон зрительных нервов доходит до передних бугров четырехолмия и уже отсюда только направляется к психооптическим центрам серой корки полушарий мозга. В согласии с этим стоит тот факт, что удаление глаза у молодого животного вызывает атрофию переднего бугра четырехолмия противоположной стороны тела. Кроме того, становится понятным, что лягушки с вырезанными полушариями все же поворачиваются своими мордочками к свету; все дело объясняется, конечно, световым рефлексом с глаза через зрительные чертоги на мышцы тела (зрительные чертоги у птиц, лягушкоподобных, земноводных и рыб являются гомологами четырехолмий у млекопитающих). В четырехолмиях заложены рефлекторные центры для движений глазного яблока и зрачка. Центры эти связаны между собою таким образом, что когда глаза сводятся книзу и кнутри, как это бывает при смотрении вблизи, то зрачки суживаются, и наоборот, они расширяются при параллельном положении глазных осей. Смотря по месту электрического раздражения четырехолмий, получается сочетанное движение глаз и т. д. Центры этих ассоциированных движений лежат, по-видимому, под четырехолмиями в передней части дна Сильвиева водопровода (Гензен, Фёлькерс). Кроме движений глаз, задние бугры четырехолмий играют, по-видимому, роль в координации движений членов, необходимых для сохранения равновесия тела, и в этом деле принимают участие особенно задние бугры четырехолмий. По Феррье, задние же бугры четырехолмий принимают участие в выражении эмоций, так как глубокое раздражение этого органа вызывает сильные крики и общие движения животного. Сопровождающие это раздражение движения желудка, кишок и пузыря, а также повышение кровяного давления и изменения в дыхании и т. д. являются последствием возбуждения через четырехолмия центров, заложенных в других частях мозга.
5) Полосатые тела и зрительные бугры, называемые большими узлами основания мозга, являются прежде всего главными путями сообщения между полушариями большого мозга и мозговыми ножками. Только некоторые волокна мозговых ножек направляются к серым извилинам полушарий мозга, не заходя в эти узлы и не заканчиваясь в их центрах, тогда как нижние, или передние, волокна (crusta) мозговой ножки проходят в полосатые тела, а верхние, или задние, волокна (tegmentum) мозговой ножки в зрительные бугры и уж оттуда волокна направляются к извилинам больших полушарий. Так как нам известно уже, что в tegmentum мозговой ножки проходят чувствующие пути, а в crusta — двигательные, то ясно, что функции полосатых тел должны быть по преимуществу двигательными, тогда как — зрительных бугров — чувствующими. В пользу того, что полосатое тело участвует в передаче и в развитии центробежных двигательных импульсов, распространяющихся от извилин Г. мозга к периферии, а зрительные бугры подобным же образом заведуют передачей и выработкой центростремительных чувствующих импульсов, говорят многие клинические факты, доказывающие, что повреждение или разрушение одного полосатого тела ведет к гемиплегии, т. е. параличу противоположной стороны тела, а разрушение одного зрительного бугра — к гемианестезии, т. е. потере в ней сознательной чувствительности. Одновременное же разрушение полосатого тела и зрительных бугров на одной стороне ведет к гемиплегии и гемианестезии противоположной стороны тела (Карпентер, Тодд, Шарко и др.). Повреждение полосатого тела может повести — согласно с двигательными функциями его - к судорогам в противоположной половине тела. Феррье, возбуждая электрическим током полосатое тело, вызывал у животных судороги в противоположной половине тела, и эта последняя сильно изгибалась в форме дуги; раздражение же зрительных бугров этого явления не давало. С этим выводом вполне согласуются опыты Карвиля и Дюре, но расходятся наблюдения Нотнагеля над функциями чечевичных ядер (nuclei lenticulares), составляющих внежелудочковые части полосатых тел. Впрочем, наблюдения эти не могут поколебать достоверности вышеуказанных экспериментальных и клинических фактов. В дополнение укажем на то, что раздражение полосатого тела впрыскиваемой в него хромовой кислотой вызывает на кроликах насильственное движение бега. Здесь, по Нотнагелю, лежит беговой узел. К заключениям, сделанным на основании опытов с раздражением или разрушением тех или других участков мозга, следует относиться с большой осторожностью ввиду того, что раздражение может не только вызывать движение, но и подавлять существующее, а повреждения известных частей мозга могут выключить те или другие функции не только вследствие разрушения определенных центров и проводников, но также и вследствие угнетения остающихся нервных механизмов.
6) Полушария мозга являются органами развития сознательных ощущений, произвольных движений и мысли (разум, чувства инстинкты), к этому выводу приводят нас как клинические наблюдения на людях и опыты на животных, так и сравнительно-анатомические данные. Животные, будучи лишены полушарий мозга, утрачивают (Флуранс) разум, сознательное чувство и волю и обращаются как бы в сложную машину, приводимую в действие раздражениями из внешнего мира или из глубины органов. Голубь, напр., лишенный полушарий мозга, способен благодаря остальным частям головного мозга сохранять равновесие тела, ходить, летать глотать, если ему вводят пищу в глотку, и т. д., но без внешних раздражений не производит ни одного из этих актов, стоит неподвижно как истукан и мог бы околеть от голоду, сидя в мешке с овсом. В начале этого столетия Галль в своей пресловутой "Système phrènologique" приписывал каждую отдельную душевную способность отдельным строго локализированным частям полуш. мозга, и таким образом им отличались мозговые бугры, или выпуклости, сильного ума, воли, хитрости, таланта к искусствам и т. д. Эта ни на чем серьезно не основанная система мозговых локализаций просуществовала недолго. Флуранс, опираясь на опыты на животных, утверждал, что полушария мозга в функциональном отношении представляются во всех частях своих совершенно однородными и послойное удаление вещества полушария мозга ведет не к исключению той или другой функции, а к равномерному ослаблению и угасанию всех психических и волевых актов безразлично; только Бульо и Андраль, опираясь на патологические наблюдения, в которых местные поражения определенных частей полуш. мозга вели к местным же параличам, полагали, что в полушариях мозга должны существовать топографически отдельные двигательные центры, соответствующие движениям конечностей, лица и т. д. Ввиду того, однако, что обширные повреждения полушарий мозга не сопровождались иногда при жизни больных никакими двигательными расстройствами или изменениями чувствительности, теория локализации функций в полушариях мозга не имела в начале успеха. Только последующие наблюдения Брока над частым совпадением потери речи у человека (афазии) с поражением третьей левой мозговой извилины и Гёглингс Джексона — над эпилептовидными односторонними и локализированными судорогами в конечностях, обусловленными местным раздражением полушарий мозга, продолжали все же служить надежной опорой теории локализации функций в полушариях мозга, которая в опытах Фритча и Гитцига, Феррьё, Карвилля, Дюре, Франка, Зольтманна, Тарханова, Бехтерева, Гарслея, Мунка и др. нашла полное экспериментальное подтверждение. Опытами этими доказано, что на поверхности серой коры полушарий мозга у млекопитающих животных существуют, с одной стороны, отдельные ограниченные двигательные площади, раздражение которых вызывает местные движения всегда в одной и той же группе мышц противоположной половины тела, а с другой — площади или центры, чувствующие для восприятия ощущений. Факты эти благодаря клиническим наблюдениям Шарко и его учеников нашли блестящее подтверждение, и ныне большинство физиологов и клиницистов, несмотря на возражения Шиффа, Броун-Секара, Германна и др., признают за твердо установленную теорию локализации функций в серой коре полушарий мозга. Электрическим раздражением различных точек обнаженной поверхности полушарий мозга у высших животных, собак, обезьян и др. доказано существование в извилинах мозга отдельных двигательных площадей для различных мышечных групп. Так, найдены отдельные двигательные центры для задних и передних конечностей, лица, языка и т. д. У обезьян двигательные участки серой коры полушарий мозга расположены возле Роландовой борозды в лобной, восходящей теменной извилинах и в парацентральной дольке. Вырезывание или повреждение этих площадей сопровождается потерей произвольного движения в соответствующих этим площадям мышечных группах противоположной стороны тела. То же приблизительно наблюдается и на человеке при разрушениях соответствующих площадей серой коры полуш. мозга кровоизлияниями (при апоплексиях), опухолями, процессами перерождения, размягчения мозговой ткани и т. д. Клинические наблюдения над различного рода односторонними местными параличами, подкрепляемые посмертным вскрытием и точным определением поврежденных во время жизни участков мозга, послужила в руках Шарко и его школы главным основанием для локализации двигательных площадей полушарий мозга у человека. В этом отношении существует большая аналогия топографии двигательных центров обезьяны и человека. В двух случаях, где исследователи непозволительно дерзнули раздражить электрическим током обнаженные вследствие изъязвления черепных костей извилины полушарий мозга у человека, они убедились, что двигательные площади заложены действительно в восходящей теменной извилине и у Роландовой борозды. В местах корки, обладающих двигательными функциями, заложены нервные центры большой величины, вроде двигательных центров спинного мозга, названные гигантскими клетками (Бетц и Мержеевский). У новорожденных слепорождающихся животных (щенки, котята, кролята и т. д.) двигательные площади серой коры полушарий мозга совершенно не развиты (Зольтманн, Тарханов) и только на 10-15 день от рождения появляются двигательные участки в корке полушарий мозга. Животные же, рождающиеся зрячими и способными сразу ходить, бегать, напр. морские свинки, уже обладают с момента появления на свет и даже в последние дни внутриутробной жизни готовыми двигательными центрами серой коры полушарий мозга, как на это указал впервые Тарханов; им же указано было, что у новорожденных животных без двигательных площадей в серой коре нет еще и гигантских центров и волокна внутренней капсулы лишены миелина, тогда как у новорожденных морских свинок эти центры уже на лицо и нервные волокна капсулы окружены миелиновым футляром. Найденные разницы Прейер распространяет на всех животных млекопитающих, относящихся, с одной стороны, к группе рождающихся зрячими и способными к координированным движениям, а с другой — рождающихся слепыми и неспособными к правильному передвижению членов.
Доказано, прежде всего Брока, а затем подтверждено Шарко и его школой, что в третьей лобной извилине левого полушария мозга локализируются центры речи; потеря речи у человека (афазия) наступает при повреждениях, кровоизлияниях именно в это место левого полушария мозга. Согласно с этим, афазия чаще всего сопровождается правой гемиплегией, т. е. параличом правой половины тела, так как эта половина заведуется левым полушарием. Мы по отношению к речи представляемся "левомозгими" (left brained), как по отношению к большинству телесных движений "праворукими", и согласно с этим левое полушарие мозга представляется большего объема и с большими извилинами, чем правое полушарие мозга. Замечательно, что левши, пользующиеся по преимуществу правым полушарием при производстве ручных работ, употребляют то же правое полушарие и для акта речи, что подтверждается тем, что афазия у левшей получается при поражениях лобной извилины правого полушария; Вильям Огль нашел три случая, в которых повреждения локализировались в правом полушарии и все три субъекта были левшами. Кроме двигательных площадей, в серой корке полушарий мозга находятся и чувствующие площади, где заложены чувствующие центры высшего порядка — психического зрения, слуха, осязания и т. д., т. е. центры, возбуждение которых ведет к возникновению специальных зрительных, слуховых и др. ощущений, осложненных сознательным элементом. Вырезывание этих чувствующих площадей полушарий мозга ведет к психической слепоте, к психической глухоте и т. д. (Феррьё и Мунк), т. е. животные, хотя и способные после этого реагировать на свет, звук, обонятельные, вкусовые и осязательные раздражения, делают это рефлекторно, не связывая с этими впечатлениями никаких определенных представлений — собаки не узнают при этом, напр., своего хозяина, не понимают угрожающей им плетки или призывных звуков свистка и т. д. У обезьян, по мнению Феррьё, Мунка, Гарслея, легко удалением определенных участков серой коры сделать животное психически слепым не только на один глаз, но на одну из половин его, психически глухим на одно ухо и т. д. Таким образом составлена Феррьё и Мунком подробная топографическая карта распределения в серой коре полуш. мозга различных психочувствующих и психомоторных центров. Зрительная площадь локализируется в затылоч. долях, слуховая — в височных долях, осязательная — в теменных долях, обонятельная — в gyrus fornicatus, а вкусовая — gyrus uncunatis. Бек, опираясь на то, что всякая возбужденная точка мозга или нерва делается сравнительно с другой, покоящейся точкой электроотрицательной, прикладывая электроды от чувствительного гальванометра к различным извилинам полушарий мозга и раздражая последовательно глаз и слух, доказал, что при раздражении светом электроотрицательное напряжение возникает в тех местах полушарий мозга, где локализируются психозрительные центры, а при раздражении слуха звуком — в местах локализации психослуховых центров. Этим доставлялось новое доказательство верности теории локализации чувствующих функций в серой коре полушария мозга. Гольц, опираясь на свои опыты с удалением у собак серой коры полушарий мозга и последующим наблюдением их, утверждает, что животные при этом не представляют никаких стойких параличей или анестезий или утраты высших чувств, а просто тупеют и тем более, чем больше удалено серого вещества. Тем не менее, ему все же пришлось признать, что передние части корки полуш. мозга стоят в более близком отношении к движениям тела, нежели задние части. Если Феррье, Мунк, Гарслей уже своей детальной локализацией кортикальных центров перешли те разумные границы, в пределах которых можно было бы отстаивать учение о локализации функций в полушариях мозга, то Гольц грешит, конечно, смелым переносом явлений с мозга собаки на мозг обезьяны и человека, относительно которых он не имеет никакого личного опыта; отрицать локализацию двигательных и чувствующих функций в коре полушарий мозга, принимая в расчет все вышесказанное, невозможно, и учение это имеет уже ныне большое практическое значение как в смысле диагностирования места поражения полушарий мозга при различных параличах, анестезиях, контрактурах или конвульсиях, так и хирургического лечения. От центров двигательных и чувствующих коры полушарий мозга идут внутрь лучисто нервные волокна, составляющие лучистую корону (corona radiata); часть этих волокон направляется к большим узлам основания Г. мозга — полосатому телу и зрительному бугру; другая же часть, суживаясь в более узкий пучок, прямо переходит во внутреннюю капсулу (capsula interna), направляющуюся далее в мозговые ножки и через них в продолговатый и спинной мозг. В состав внутренней капсулы входят, кроме этих кортикальных волокон, еще и волокна, идущие от больших узлов основания мозга. Опыты показали, что разрушение передней трети внутренней капсулы, происходящей от прелобной дольки, не изменяет ни движения, ни чувствительности; разрушение средней трети внутренней капсулы, находящейся в связи с лобно-теменными извилинами, обусловливает стойкие двигательные параличи в противоположной половине тела с последовательным перерождением нервных путей и стойкими контрактурами и что разрушение задней трети внутренней капсулы, волокна которой происходят от височно-затылочных областей полушарий мозга, вызывает параличи различных форм чувствительности, т. е. анестезии.
Полушария мозга являются, кроме всего сказанного, еще и местом психической деятельности, скрывающейся в форме чувств, составляющих аффективную сторону душевной жизни (настроений); в форме представлений, обнимающих сферу разума, воспоминания и фантазии; к этой области относятся: содержание представлений, формальное течение их, скорость их течения, ассоциация и воспроизведение их; и в форме стремлений (к пище, половая страсть, деятельность воли). Сознание является суммой имеющихся в определенный момент представлений, а самосознание есть сознание самим субъектом протекающей в нем самом работы представлений. Большинство заученных движений в бодрствующем состоянии производится средними частями Г. мозга, без всякого участия сознания. Во время глубокого сна сознание отсутствует, подобно тому, как оно отсутствует вполне и у животных без полушарий мозга, — все внешние проявления разумности, понятливости, выбора, инициативы и воли совершенно отсутствуют у таких животных, и такое состояние является стойким и постоянным. В психич. актах различают след. моменты: 1) Доставка к мозгу по центростремительным, или чувствующим, нервам каких-нибудь возбуждений, вызванных внешними или внутренними раздражителями; 2) акт восприятия их в сознании, чему способствует особое возбужденное состояние в психическом органе, именуемое вниманием; 3) внутреннее созерцание, соответствующее действительному изучаемому явлению и основанное на вызове образа воспоминания, данного уже предварительным психическим развитием, и, наконец, 4) волевой импульс, т. е. перенос возбуждения на один или на группу двигательных центров, а через них на 5) центробежный нерв и на мышцы, приводимые в сокращение. Из всех этих звеньев психического акта явления под рубриками 2), 3) и 4) протекают прямо в серой коре полушарий мозга, а потому разрушение ее ведет к прекращению психической жизни. Что касается локализации различных психических способностей — чувств, представлений, воли — по различным участкам полушарий мозга, то об этом решительно ничего нельзя сказать достоверного; полагают только, что органы воли, а быть может и мышления, лежат преимущественно в лобных долях полушарий мозга; но предположения эти требуют дальнейших фактических подтверждений. Существует очень явное и оригинальное доказательство на лягушках связи психических функций с полушариями мозга, основанное на вызове у этих животных галлюцинаций и иллюзий при исчезании хлороформного наркоза. В известных стадиях проходящего наркоза у них последовательно появляются: фаза каталептическая, когда они сосредоточены как бы на каких-то ощущениях; фаза галлюцинаций и иллюзий — сказывающаяся ловлей не существующих вокруг них предметов, насекомых, сопровождаемая хватательными движениями челюсти, набрасыванием на окружающих и эротическим стремлением к обхватыванию самок; эта агрессивная фаза заменяется манией преследования, во время которой лягушки то и делают, что телодвижениями защищаются от не существующих вокруг них врагов и под конец обращаются в настоящее нормальное бегство от человека, когда наркоз уже совершенно прошел. Таким образом, в ряде иллюзий и галлюцинаций выражаются все существенные стороны жизни лягушки. Замечательно, что все это наблюдается только при наличности полушарий мозга (Тарханов).
Несомненно, что богатство психического мира зависит не столько от объема полушарий мозга, сколько от богатства их серой коры мозговыми извилинами (увеличивающего массу вещества, специально содержащего нервные центры), от обилия так называемых ассоциационных волокон, связывающих между собою извилины одного и того же полушария, от обилия комиссурных волокон, связывающих оба полушария между собою. По крайней мере, мозг идиотов как раз скуден только что указанными анатомическими особенностями, составляющими атрибут мозга, способного к нормальной психической деятельности.
Приводимые обыкновенно таблицы веса Г. мозга различных великих людей, доказывающие будто бы известную параллельность между силой ума или таланта и объемом мозга, вовсе ничего не доказывают (см. Головной мозг человека). Психические функции полушарий мозга находятся в теснейшей зависимости от кровообращения, и прекращение его, хотя бы очень кратковременное, при остановках сердца вследствие испуга, различных аффектов и т. д. вызывает обморок, обыкновенно проходящий при возобновлении нормальной деятельности сердца. Многие наркотические вещества, хлороформ, эфир, алкоголь, морфий, хлоралгидрат и т. д., примененные в больших дозах, уничтожают на время раздражительность серой коры полушарий мозга, психомоторные и психочувствующие центры переходят в недеятельное состояние, и сознание исчезает (Феррьё, Мунк, Фритч и Гитциг, Гейденгайн и др.). Этим и пользуются обыкновенно при производстве болезненных операций. Раздражительность психомоторных площадей полушарий мозга резко падает и во время нормального сна животных, как это легко констатировать на спящих щенках со вскрытым Г. мозгом (Тарханов). Серая кора полушарий мозга орошается более богатой сетью кровеносных сосудов, нежели белое вещество их; во время нормального и искусственного сна сосуды мозга беднее кровью, нежели во время бодрствования. Прилив артериальной крови к мозгу регулируется артериальным кругом Виллизия (Circulus аr teriosus Willisii), прекрасно выравнивающим, лучше чем где-либо, резкие колебания артериального давления в черепной полости. Если перевязать обе сонные и обе позвоночные артерии, то всякий прилив крови к мозгу прекращается и тотчас наступают сильные эпилептовидные судороги с потерей сознания, т. е. полный эпилептический припадок. При вскрытой черепной крышке после трепанаций или после разъедания черепных костей различными язвами и т. д., а также и на родничках у детей легко заметить пульсационные и дыхательные колебания объема мозга. При помощи регистрирующих аппаратов легко записать эти колебания мозга, причем оказывается, что маленькие приподнятия мозга соответствуют приливу крови к мозгу при каждом сердечном сокращении, а маленькие опускания — периоду расслабления сердца, тогда как на больших колебаниях мозга максимумы приподнятия соответствуют акту выдыхания, а максимумы опускания — акту вдыхания. — Колебания объема мозга являются, таким образом, отражениями тех изменений в мозговом кровообращении, которые вызываются периодической деятельностью сердца и дыхательного аппарата. — Важно напомнить здесь, что все формы возбуждения органов чувств и психической деятельности человека сопровождаются увеличением объема мозга на счет усиленного прилива к нему крови, т. е. к деятельному мозгу всегда усиленно приливает кровь — это констатировано как на человеке, так и на животных. Во время же сна объем мозга падает, как это легко видеть на родничках у детей. При замкнутой со всех сторон черепной крышке такие приливы и отливы крови из мозга мыслимы только благодаря свободному переливу цереброспинальной жидкости из Г. мозга в спинной; при усиленном приливе крови к мозгу цереброспинальная жидкость переливается в спинной мозг, и наоборот, при сжатии кровеносных сосудов она вновь возвращается в черепную полость. В полушариях мозга лежат также центры, возбуждение кот. отражается и на сердцебиениях, на кровеносных сосудах, дыхании, слюноотделении и на многих др. процессах растительной жизни организмов, но о них речь будет в местах, касающихся описания этих процессов.
И. Тарханов.
|
Page was updated:Tuesday, 11-Sep-2012 18:15:05 MSK |