| [ начало ] | [ К ] |
Клеточка
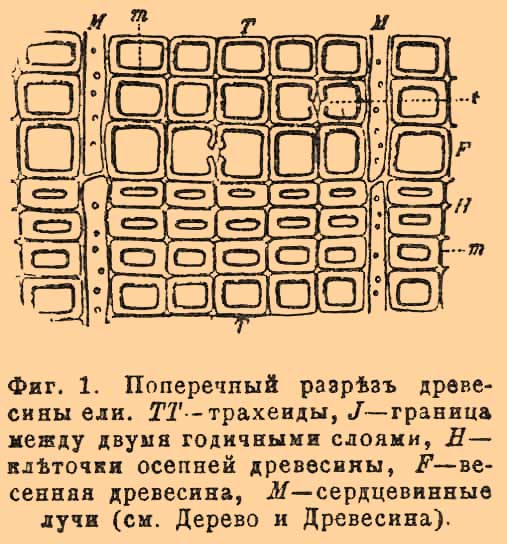
(cellula). — Рассматривая под микроскопом, даже при слабом увеличении, тонкие пластинки, вырезанные из всевозможных частей растений и животных, можно скоро убедиться, что они не состоят из однородной массы, а слеплены из бесчисленного множества чрезвычайно мелких частиц. Особенно ясно это у взрослых растений, где каждая из таких частиц представляется микроскопически мелкой камерой, отграниченной от соседних с ней таких же камер посредством перегородок (фиг. 1).
Фиг. 1. Поперечный разрез древесины ели. TT — трахеиды, J — граница между двумя годичными слоями, H — клеточки осенней древесины, F — весенняя древесина, M — сердцевинные лучи. (см. Дерево и Древесина.)
Поэтому старинные знаменитые ученые, впервые изучавшие и писавшие о внутреннем строении растений, говорили, что растение построено, как пчелиный сот, — из ячеек или клеточек, из камер, отделенных друг от друга подобно тому, как отделены ячейки или клетки сотов восковыми перегородками. Название К. и до сих пор осталось за этими элементарными органами или организованными элементами растений и животных. Изучение внутреннего строения организмов, продолжавшееся два столетия, привело окончательно к установлению того факта, что самая важнейшая составная часть К., а именно протоплазма с ядром, в ней заключенным, свойственна всем живым К. без исключения, будут ли они растительные или животные. В этом смысле и говорят, что все организмы состоят из К. Это же побуждает некоторых ученых (например, Оскара Гертвига) определять К. как комочек протоплазмы, заключающий внутри себя особую форменную составную часть — ядро (nucleus). Громадное большинство взрослых растительных К., однако же, отличается еще тем, что их протоплазма одета снаружи особой оболочкой, состоящей из вещества иного физического и химического строения, чем протоплазма, которая во взрослых растительных клеточках занимает далеко не все пространство внутри оболочки, так как оно содержит еще водянистый сок, называемый клеточным. К. животных также нередко имеют наружную оболочку, но она не столь резко, как у растений, отличается от протоплазмы и в большинстве случаев даже от нее не вполне обособлена, являясь только наружным отверделым ее слоем. Итак, будучи сходны и даже одинаковы между собой в главной составной своей части, К. растительные и животные тем не менее отличаются между собой указанными выше чертами, а отчасти и способами своего размножения. Тем не менее протоплазма с ядром и с включенными в нее крупчатыми, капельножидкими и другими телами представляет собой обособленное целое, так называемый протопласт, а у животных, как уже сказано, в большинстве случаев являет собой и всю К. У растений К., хотя и редко, но тоже в некоторых определенных случаях состоит из одного только протопласта, и тогда она называется голой и вполне сходна с большинством К. животных. Сходство это особенно разительно на первых степенях развития, ибо яйцо, коим начинается каждое животное и растение, есть не что иное, как голая К., т. е. комочек протоплазмы с ядром. Исключение составляют только споры тех грибов, у которых до сих пор не найдено половых органов, но и тут протопласт споры соответствует яйцу высших растений. Голые клеточки нередко представляют весь организм, и тогда он, очевидно, является одноклеточным. Таковы корненожки (фиг. 2), таковы зооспоры или бродяжки многих низших растений: водорослей и грибов.
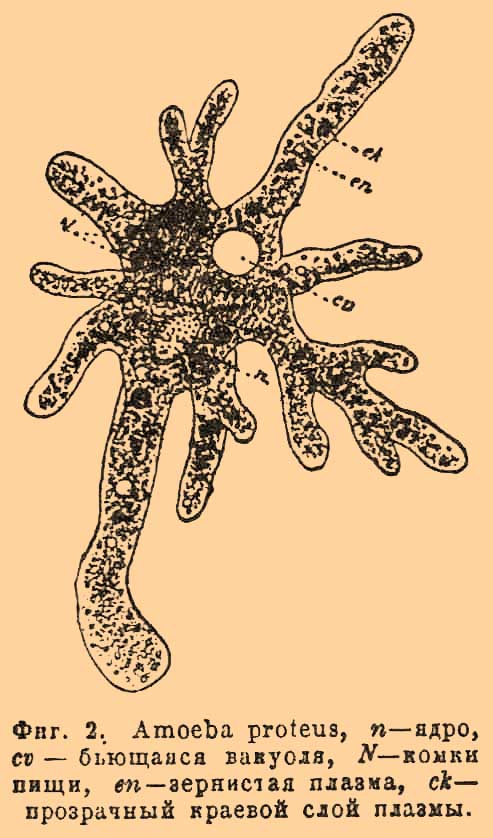
Фиг. 2. Amoeba proteus. n — ядро, cv — бьющаяся вакуоль, N — комки пищи, en — зернистая плазма, ck — прозрачный краевой слой плазмы.
У слизистых грибов или миксомицетов споры выпускают комочки протоплазмы (фиг. 2), которые имеют значение бродяжек, но вскоре теряют первоначальную форму, принимая форму амёб (см.) из корненожек. Эти амёбообразные бродяжки даренные, как и корненожки, движением, соединяются между собой (фиг. 3), сливаются и образуют одну крупную, видную невооруженным глазом сетчатую массу, так называемый плазмодий, который представляет собой одну очень большую голую К.
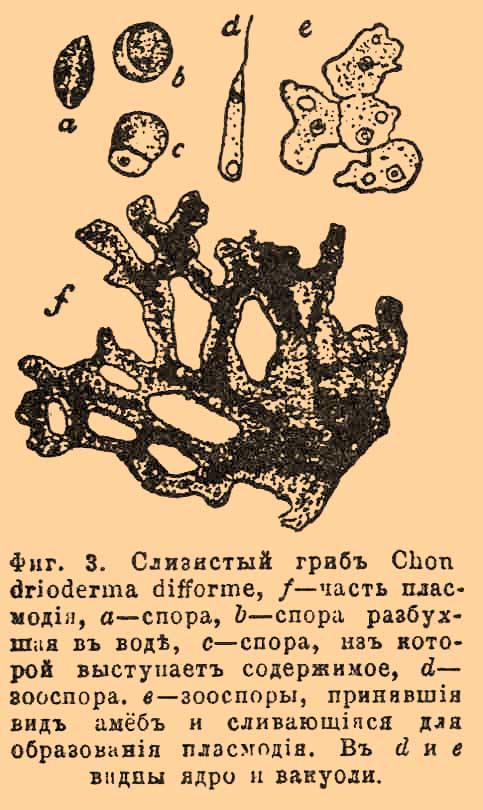
Фиг. 3. Слизистый гриб Chon drioderma difforme, f — часть плазмодия, a — спора, b — спора, разбухшая в воде, c — спора из которой выступает содержимое, d — зооспора, e — зооспоры, принявшие вид амеб и сливающиеся для образования плазмодия. В d и e видны ядро и вакуоли.
В таком большом объеме голая протоплазма только у слизистых грибов и появляется, а потому свойства протоплазмы всего удобнее изучаются именно на плазмодии. Тут, как и во всех остальных случаях, протоплазма представляется почти всегда прозрачной слизью, не смешивающейся с водой и сильнее ее преломляющей свет, так что в воде она всегда ясно различается. В этой слизи или вязкой жидкости, как ее называют новейшие авторы, содержится по большей части огромное число мельчайших крупинок, так называемых микросом. В ней же заключено одно или несколько ядер, а нередко и мелкие полости, наполненные прозрачным соком, так называемые вакуоли, хотя тут о настоящей пустоте (vacuum) не может быть и речи. Крупицы бывают весьма различной величины. Их не бывает только в самых наружных слоях протоплазмы; поэтому края плазмодия и всякого комка протоплазмы совершенно прозрачны, нередко даже плотнее остальной массы, и принимались прежде за особый кожистый слой, хотя настоящей кожи тут нет. В молодых растительных тканях протоплазма сплошь наполняет К. (фиг. 4 А), но по мере их разрастания образуются вакуоли, которые сливаются между собой, раздробляют протоплазму на нити и оттесняют ее к поверхности, так что она образует подкладку под оболочкой К., придерживая около этой оболочки ядро (фиг. 4 С).

Фиг. 4. Продольные разрезы из корневой коры Fritillaria imperialis, A — совершенно молодые К., еще не содержащие сока; B — такие же клеточки, но старые. Образовались вакуоли s, наполненные соком, p — плазма; C — такие же К., но еще старше; S — сок, kk — ядра с ядрышками, xy — ядро разбухшее, p — плазма, h — оболочка.
Если же ядро остается в середине К., то нити протоплазмы идут к нему от стенок и оно все-таки более или менее окружено протоплазмой. Для открытия постенной, иногда едва заметной протоплазмы пользуются явлением, называемым плазмолизом (фиг. 5).
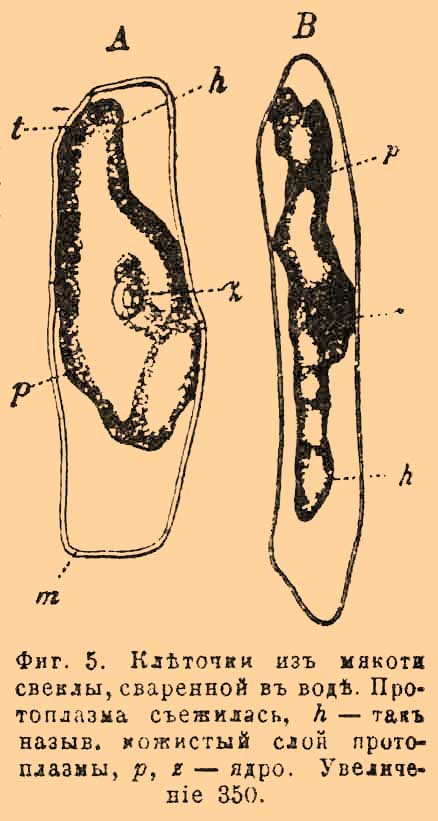
Фиг. 5. Клеточки из мякоти свеклы, сваренной в воде. Протоплазма съежилась, h — так называемый кожистый слой протоплазмы, p, z — ядро. Увеличение 350.
Если погрузить живую растительную К. в слабый раствор сахара, то протоплазма стягивается в виде туманного мешочка внутрь К., отходя от ее стенок. Плазмолиз основан на том, что растворы солей или сахара, проникая диасмотически через внешнюю оболочку К., не проникают сквозь протоплазму. В новейшее время на очереди вопрос о тончайшем строении протоплазмы (фиг. 6).
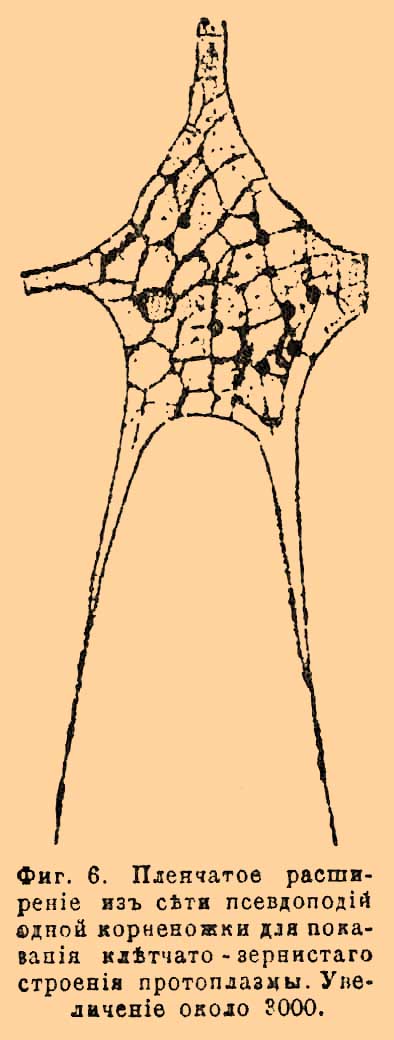
Фиг. 6. Пленчатое расширение из сети псевдоподий одной корненожки, для показа клетчато-зернистого строения протоплазмы. Увеличение около 3000.
На этот счет имеется несколько теорий, но ни одна из них не получила еще права гражданства в науке. До сих пор выяснилось лишь то обстоятельство, что протоплазма есть смесь многих химически и физически различных веществ, но в точности не известно, имеется ли какая-нибудь законность в их взаимном расположении. Само химическое строение протоплазмы могло быть изучено до сих пор только в общих чертах. Химический анализ ее затруднен, во-первых, тем, что ее невозможно получить в достаточном количестве, а затем необыкновенной сложностью ее состава и многочисленными включениями в нее, удаление которых почти невозможно. Несомненно, однако же, то, что во всякой протоплазме преобладают белковые или, как их многие называют, протеиновые тела. Белки представляют сложнейшие в химическом отношении вещества. В состав их входят углерод, водород, кислород, азот и сера. Но кроме этих тел, протоплазма заключает еще некоторые соли, в которых открываются, а именно в золе после сжигания: калий, натрий, магний, кальций, железо, хлор, сера и фосфор. Все это найдено у слизистого гриба Fumago varians, но, по всей вероятности, перечисленные тела находятся, хотя отчасти, и в протоплазме всех организмов (см. Белковые вещества). Кроме белковых тел и включений, протоплазма содержит много воды, так, например, у Fumago varians найдено 71,6% воды, у других меньше, но все же значительно. Высыхая, протоплазма может получать почти свойство воска и в этом виде сохраняться довольно долго, не теряя жизненности. При смачивании такой протоплазмы она снова оживает. Другая, не менее важная часть протопласта — ядро. В настоящее время полагают, что все К., как животные, так и растительные, его содержат. В большинстве случаев каждая К. заключает только одно ядро, но имеются К. с большим их числом, например у многих грибов и водорослей (см. фиг. 7), в почечных К. по 2 ядра, в К. костяного мозга их бывает до 100 и более. Помногу ядер у многих корненожек, инфузорий и проч.
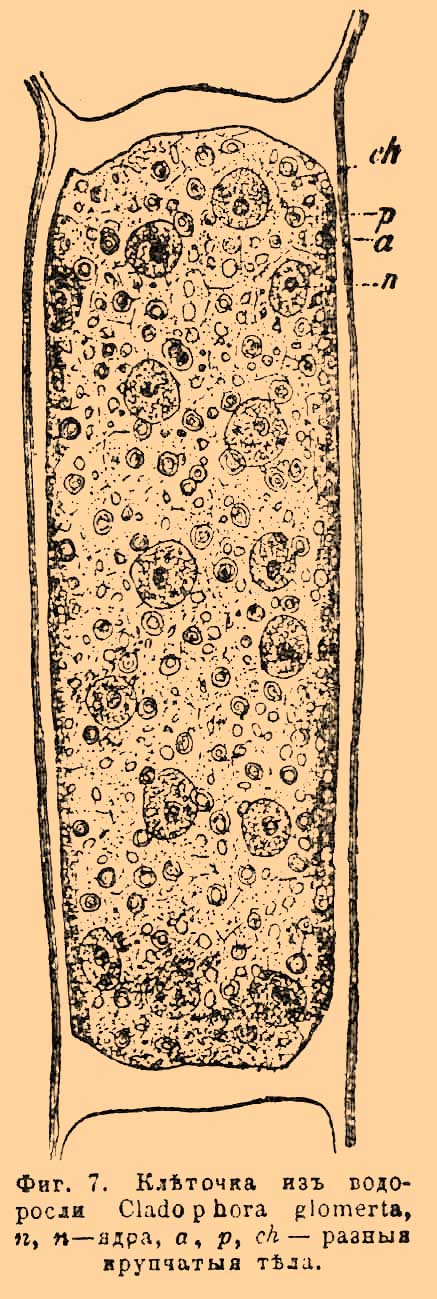
Фиг. 7. Клеточка из водоросли Cladophora glomerta, n — ядра, a, p, ch — разные крупчатые тела.
Величина ядра почти всегда микроскопически мала, но есть и видные простым глазом, в виде точек, например в незрелых яйцах амфибий и рыб. Форма по большей части овальная или шаровидная, но есть и другие формы, даже ветвистые, как у некоторых мелких рачков. Оно снабжено нередко оболочкой, особенно у растений; внутри этой оболочки заключена самая масса ядра, состоящая из нескольких веществ, между которыми обильнее других так называемый нуклеин или хроматик — белковое вещество, содержащее фосфор. Внутри ядра он принимает форму то отдельных зернышек, то тонкой сети или нитей. Другое белковое тело — парануклеин или пиренин, отличающееся своими микрохимическими реакциями от хроматика, входит в состав ядрышек, замечаемых в каждом ядре. Эти 2 тела считаются многими за важнейшие из составных частей ядра. Второстепенными его составными телами считается линин, образующий нередко сетки и перекладники в ядре. Он отличается, между прочим, своей неспособностью окрашиваться обыкновенными красильными веществами. Кроме всего этого, в ядре замечается сок, подобный тому, что имеется в протоплазме: он наполняет промежутки между построениями, состоящими из вышеназванных тел. Наконец, оболочка ядра, которую некоторые ученые (например, Страсбургер) считают за кожистый слой протоплазмы, облекающей ядро, состоит опять-таки из белкового вещества, отличного по своему физико-химическому составу от остальных. Прибавить должно, что все эти тела, подобно тем, что заключены в протоплазме, вовсе еще не исследованы точным образом и известны только в общих чертах.
Кроме ядра в К. наблюдается часто одно или 2 очень маленьких тельца, лежащих около ядра. Это так называемые центросомы; они имеют важное значение при размножении К. Найдены ныне в животных, так и в растительных К.; самостоятельность их и присутствие во всех К. без исключения еще окончательно не подтверждены, хотя это весьма вероятно. Таковы главные, основные черты протопласта, но кроме того, протоплазма содержит внутри себя разного рода включения, к числу коих прежде всего следует отнести клеточный сок, содержащий все растительные и животные К. У растений это раствор, содержащий растительные кислоты и их соли, сахар, белковые вещества и проч. Сок этот наполняет собой вакуоли, сливающиеся у растений в одну общую полость, занимающую у взрослых К. почти всю внутренность К. (см. выше). У животных образование вакуолей гораздо реже, хотя и замечается особенно у одноклеточных организмов, каковы корненожки и инфузории (фиг. 2). Из других жидких веществ в протопласте нередко попадается жир, в виде мелких капелек, сливающихся нередко в крупные, что особенно распространено у животных, попадаясь, однако же, обильно и у растений в тканях, служащих для накопления запасных питательных продуктов, каковы клубни, луковицы, маслянистые семена. В яйцеклетках животных часто накопляются более или менее твердые желточные шарики, крупицы или пластинки, представляющие смесь белковых веществ с жирами. В растительных К. из числа твердых включений особенно важное значение имеют хроматофоры или пластиды (еще иначе трофопласты) и производимые ими хлорофилл (см.) и крахмал (см.). Между еще более твердыми телами, включенными часто в протоплазму растений, следует указать еще на кристаллы (см.), кристаллоиды, алейрон и проч. Протопласт, будучи седалищем жизни, проявляет все те физиологические отправления, сумма которых составляет физическую жизнь организмов. Специальное одному протопласту, а вследствие того, одним только организмам принадлежащее свойство — есть раздражимость или раздражительность, т. е. способность реагировать на различные лады на влияния общефизических внешних сил. В связи с этим по преимуществу жизненным явлением находится движение, которое свойственно протоплазме и которое особенно ясно наблюдается на голых К. Именно в плазмодии миксомицетов замечается постоянно передвижение крупинок, а также ползучее передвижение всей массы, сопровождаемое беспрестанным выпячиванием и втягиванием тонких отрогов на краю всего плазмодия. Эти отроги вполне соответствуют тем отрогам, что вытягиваются и втягиваются амёбами (см.) и называются псевдоподиями (фиг. 2). Внутри К., снабженных оболочками, у многих растений замечается также движение крупчатой протоплазмы. Это движение наблюдается с большой ясностью в длинных листовых К. валлиснерий (см.), элодей (Elodea canadeosis) и проч. Также в волосках многих растений, например в длинных корневых волосках водокраса (см.), в волосках тычиночных нитей традесканций (фиг. 8) и проч.
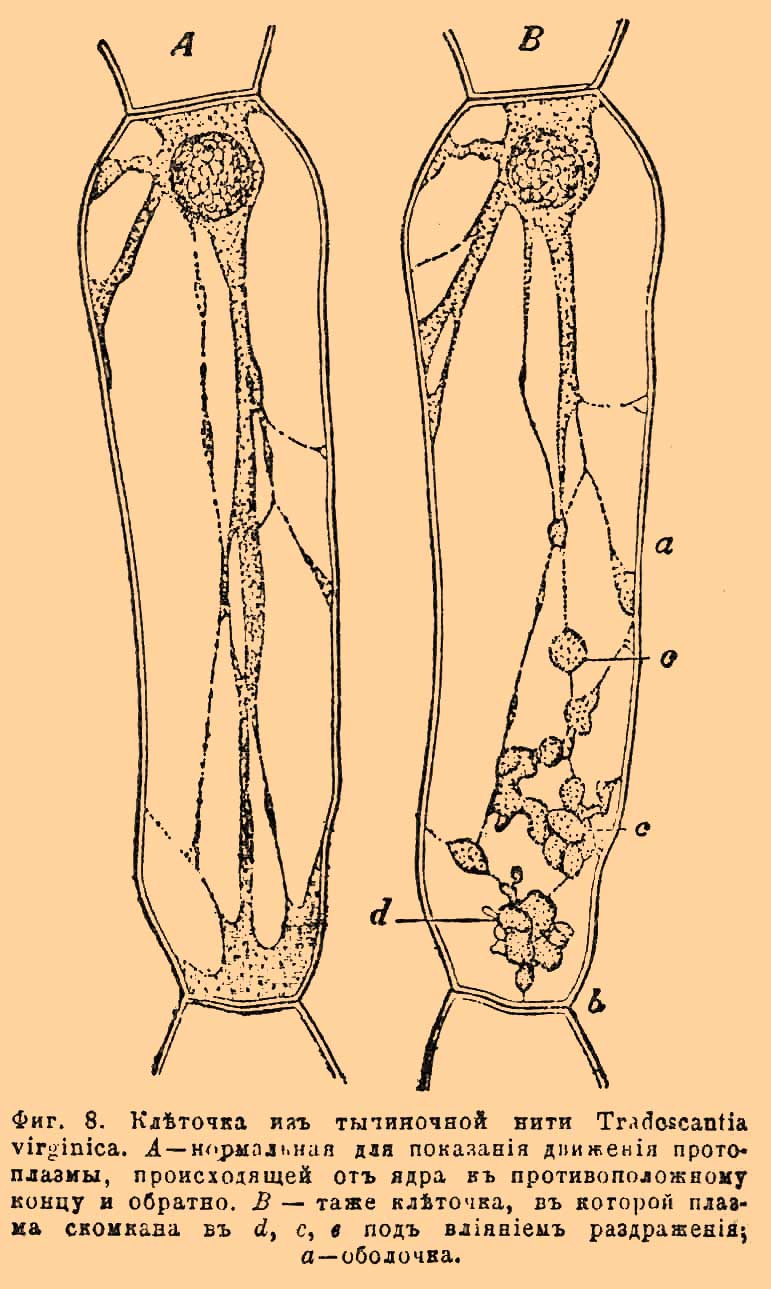
Фиг. 8. Клеточка из тычиночной нити Tradescantia virginica. A — нормальная, для показа движения протоплазмы, происходящей от ядра к противоположному концу и обратно. B — та же клеточка, в которой плазма скомкана в d, c, e под влиянием раздражения; a — оболочка.
Движение это представляется или в виде тока, идущего в постенной протоплазме от одного конца К. к другому и обратно — это так называемое вращательное движение (rotatio), или оно происходит среди нитей протоплазмы, идущих к ядру по разным меняющимся направлениям — это так называемая циркуляция. Протопласт, как мы видели, заключен у огромного большинства взрослых растительных К. в оболочку. Оболочка эта состоит из клетчатки (см.) или клетковины, т. е. из вещества группы углеводов. Она есть продукт протопласта в том смысле, что выделяется из самой протоплазмы. Поэтому некоторые ученые (например, Оскар Гертвиг) не считают ее интеральной частью К., а причисляют к межклетным веществам, куда относят и оболочки животных К., где такие попадаются, а также различного строения и свойства массы, выполняющие промежутки между К. животных. Там нередко межклетное вещество занимает гораздо больше места, чем самые К., как это замечается, например, в костях, хрящах (фиг. 9) и проч. (см. Ткани).

Фиг. 9. Хрящ. C — его кожица, b — переход к типическому хрящу (a).
Клетковинная оболочка растительных клеточек на первых степенях своего развития необыкновенно тонка, прозрачна и окрашивается йодом в голубоватый цвет, но эта способность молодой оболочки скоро исчезает, и тогда голубая окраска вызывается другими реактивами, из которых употребительнее — хлорцинковый раствор. Молодые, вновь нарастающие части растений состоят из одинаковых К., друг от друга отделенных тончайшими клетковинными оболочками; по мере возрастания клеточки оболочка разрастается во все стороны и в большинстве случаев сильно утолщается. Она сильно растяжима и упруга, способна быстро пропускать через себя, путем осмоса, воду, причем может сильно разбухать, спадаясь при испарении. Это разбухание оболочки, вследствие гидростатического давления, называется физиолого-ботаниками тургором. Оно имеет важное значение в жизни растений. Хотя оболочка в таком состоянии, а отчасти и в последующих кажется лишенной всякого строения, но в большинстве случаев в ней можно открыть полосатость по двум перекрещивающимся направлениям, что подает повод считать ее состоящей из частиц, определенным образом расположенных. Кроме того, оболочка очень часто представляет своеобразную узорчатость, зависящую от способа ее утолщения. Утолщение происходит вследствие нарастания вещества с наружной или с внутренней стороны оболочки. Наружные утолщения образуют у растений часто различные узорчатые выростки, бородавочки и т. п., как, например, на многих волосках и особенно на цветневых пылинках (см. Цветень). Утолщения изнутри представляют не только слоистость, но еще от места до места перерывы, которые при значительном утолщении оболочки представляются в виде каналов, направляющихся из внутренней полости К. к ее наружной оболочке. Отсюда и происходит узорчатость оболочки. Форма узоров определяется формой перерывов в утолщающем веществе. Если перерывы в виде незначительных округлых или длинноватых фигур, то оболочка представляется пятнистой или испещренной черточками; если перерывы в виде сетки — она сетчатая, если утолщающаяся масса в виде колец или спиральных лент, то и оболочка сетчатая, кольчатая или спиральная, и т. д. Во всяком случае перерывы представляются светлыми узорами, а самые утолщения более темными. Утолщение нередко так значительно, что полость К. почти исчезает (фиг. 10).
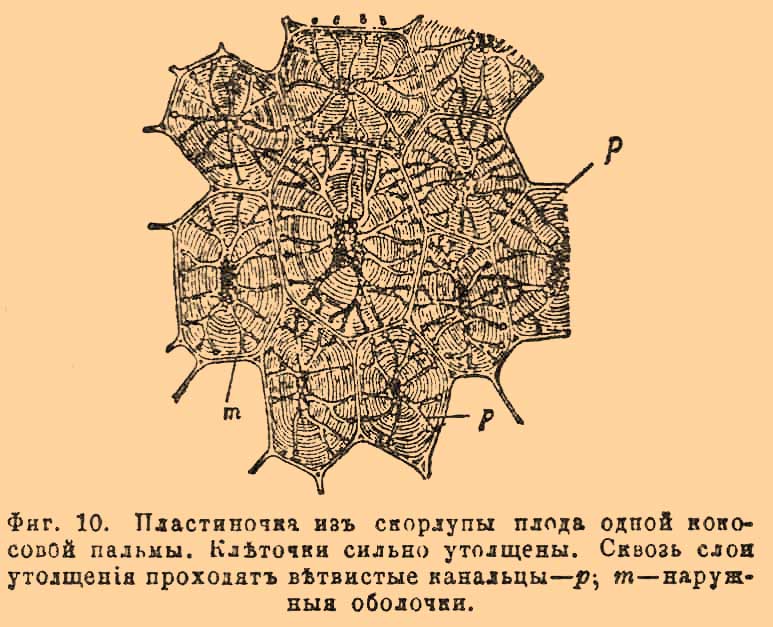
Фиг. 10. Пластиночка из скорлупы плода одной кокосовой пальмы. Клеточки сильно утолщены. Сквозь слои утолщения проходят ветвистые канальцы — p; m — наружные оболочки.
Возрастание в плоскости и утолщение объясняется двумя процессами: внедрением (интуссуссенция) новых частиц между старыми и отложением (аппозиция) нового вещества на первоначальную оболочку. Одним из этих процессов объяснит все явления возрастания нельзя, участие обоих единовременно необходимо. Общая форма клеточек различна так же, как и их величина. Свободно лежащие или рыхло соединенные К. имеют или совершенно сферическую форму, например яйцевые клеточки растений и животных, или более или менее округлую: яйцевидную, грушевидную, чечевицеобразную, как, например, кровяные шарики человека и многих животных (см. Кровь). Тесно соединенные К. бывают гранеными, чаще всего 6-гранными (в сердцевине многих растений). Кроме того, К. бывают формы звездчатой, как, например, в сердцевинной ткани многих растений (ситниковые, осоковые), ветвистой, снабженной нередко чрезвычайно тонкими и длинными ветвями, как, например, К. нервов мозга, и проч. Бывают К. длинные (мышечные), особенно у растений, где так много трубчатых круглых или плоских (хлопчатобумажник, у которого волокна, одевающие семена, составляют хлопок). Прядильные волокна из стеблей льна, конопли также состоят из длинных, толстостенных К. (см. Ткани). Что же касается до величины К., то она чрезвычайно различна. Вообще они так малы, что измеряются микромиллиметрами — μ, т. е. тысячными долями миллиметра. К. животных бывают изредка так велики, что их можно видеть простым глазом, например К. яйца многих холоднокровных животных. Вообще же величина К. животных колеблется от 4 μ до 80 и даже 200 μ в диаметре. Растительные К. только сравнительно в редких случаях бывают видны невооруженным глазом. У низших растений, а также между волосками и внутренними волокнами высших растений имеются К. весьма заметных размеров, особенно в длину. Так, длина хлопчатобумажных К. бывает от 2 до 5 см, К. льняных волокон до 4 см. Диаметр несравненно меньше: он равен 20 μ у хлопчатобумажных К. и 14 μ у льняных. Особенно велики круглые и граненые растительные К., которые бывают иногда в 300 μ, например в сердцевине, бузины. Гигантскими К. должно считать те, например, что составляют, каждая, у рода Nitela из Хар целое стеблевое колено и бывают длиной от 8 до 11 см при диаметре в 0,8 мм.
Происхождение и размножение К. Установлено, что К. происходят не иначе как от К. (omnis cellulla e cellula). Следовательно, повсюду имеются К. производящие и производные [Вместо этой давно предлагаемой мной терминологией некоторые из наших ученых продолжают употреблять выражения матерная или материнская К. вместо производящая, дочерняя — вместо производная и сестринские К. — вместо сопроизводные. Это прямо перевод с немецкого или французского и притом неправильный, ибо окончание на -ский означает принадлежность, а не качество: материнский означает принадлежащий матери, дочерний — принадлежащий дочери, а не качество К., как матери или дочери.]. Так как каждая К. может дать начало не только одной, но также 2 и большего числа, то эти единовременно появляющиеся К. называются сопроизводными. Кроме того, стараются установить правило, по которому всякое ядро тоже происходит через деление ядра — omnis nucleus e nucleo, но хотя это явление действительно чрезвычайно распространено как в животных, так и в растительных клеточках, но наблюдается и прямое образование его среди протоплазмы, например при образовании К. почкованием у растений. Во всяком случае, относительно происхождения ядра не все еще разъяснено. Как бы то ни было, но размножению К., заключающих в себе одно или несколько типических ядер, предшествует размножение ядра или ядер. Оно совершается в большинстве случаев, как это считают новейшие исследователи, посредством так называемого кариокинеза (karyokinesis) или посредством деления, называемого также сегментацией (О. Гертвиг). Этот процесс совершается в важнейших отношениях одинаково, как в животных, так и в растительных К., что и представляет главный его интерес. Вот главные его черты. Мы видели, что ядро состоит из сеточки, в которой хроматик (см. выше) появляется в виде крупинок, а линин — в виде связующих эти крупицы нитей и перекладин. Перед началом сегментации (или кариокинеза) сетка мало-помалу распутывается, все ее части соединяются в одну общую сильно извилистую нить, которая образует среди ядерного сока извилистый остов ядра (фиг. 11).
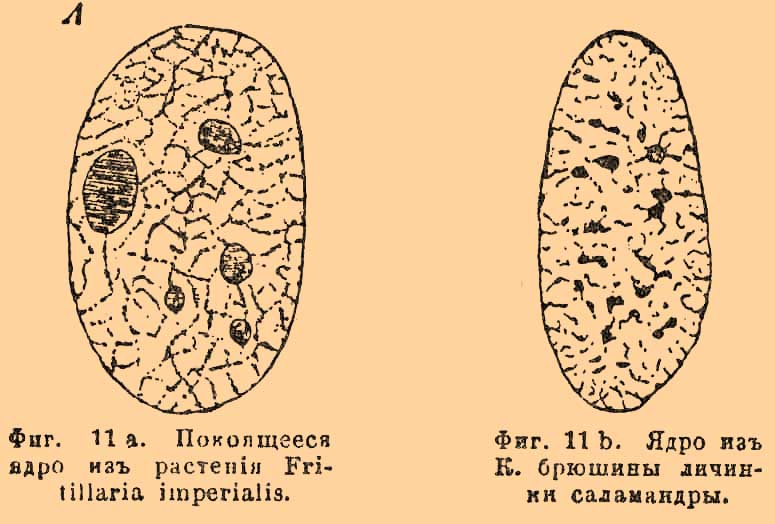
Фиг. 11 a. Покоящееся ядро из растения Fritillaria imperialis.
Фиг. 11 b. Ядро из К. брюшины личинки саламандры.
Затем извилистая нить распадается на участки, называемые ядерными сегментами. Эти сегменты получают форму крючков с одинаковой или разной длины сторонами (фиг. 12). Они собираются в серединной плоскости ядра — в его экваторе, а затем распадаются каждый вдоль на 2 половины (фиг. 12).

Фиг. 12. Схематическое изображение кариокинеза растительной или животной К.; A — ядро перед делением. B — сеть преобразовалась в извилистый шнур. C — начало сегментации. D — раздвоение. E — образование производных ядер. F — производные ядра окончательно образовались, z — образующая перегородка.
Пока это происходит, центросомы, коих обыкновенно 2 и которые находятся с боку ядра, передвигаются на полюсы ядра, один против другого, и от них начинают расходиться протоплазматические нити во все стороны, но преимущественно к середине ядра. Затем полусегменты расходятся к полюсам так, что к каждому полюсу передвигается по полусегменту каждого из разделившихся по длине сегментов. Достигнув полюсов, хроматиковые или ядерные сегменты собираются клубочками и мало-помалу образуют остовы новых производных ядер, а протоплазматические нити между тем увеличиваются в своем числе и толщине; там, где они сходятся на экваторе К., возникает накопление протоплазматических крупиц, а затем, у растительных клеток, и клеткованная перегородка. Весь процесс заканчивается не только разделением ядра, но и разделением всей К. В сравнительно редких случаях ядро разделяется с помощью простого перетягивания. Новейшие ученые полагают, что размножение К. делением всегда начинается сегментацией ядра; следует, однако же, заметить, что наблюдения в этом отношении коснулись пока весьма небольшого числа растений. Кроме деления К. надвое, может происходить образование одной производной из всего протопласта данной К. — тут, без сомнения, не может быть речи о кариокинезе. Так образуются, например, подвижные споры или бродяжки некоторых водорослей (см.) и проч. Размножение К. может совершаться также почкованием, например у бродильных грибков (см. Грибы и Размножение растений).
История. До 1675 г. не имели никакого понятия о К. и о внутреннем строении растений, но именно в этом году вышли в Лондоне обширные исследования итальянского ученого Марчелло Мальпиги и английского — Неэмии Грю. Этими учеными открыты были, а именно у растений, не только К., но и элементарные органы, из них происходящие. Эти первые наследования долго оставались без продолжения и без развития в значительной степени оттого, что и самый инструмент, т. е. микроскоп, без которого немыслимо изучение элементарного строения организмов, долго оставался неусовершенствованным. Мало-помалу в конце XVIII и в начале нашего столетия, трудами французских (Тюрпен, Бриссо-Мирбель, Распаль, Дютроше) и немецких ученых (Тревирануса, особенно Гуго Моля, Мейена), познание морфологии и отчасти физиологии К., преимущественно растительной, подвинулось вперед. Менее успешно шли исследования тончайшего строения животного организма, но уже в тридцатых годах нашего столетия разными учеными, каковы, например, Генле, И. Миллер и проч., животные клеточки сравнивались с растительными. Важнейшим шагом вперед на пути исследования К. следует считать окончательное установление полной гомологии, сходственности в основных чертах растительной и животной К. Эта мысль уже высказывалась Распалем и Дютроше, но фактически доказана и введена в науку Шлейденом (1838) и Шванном (1839). Первый из этих ученых, пользуясь открытием клеточного ядра Робертом Брауном (1833), доказал очень обширное его распространение и важное значение в размножении К.; второй, в своих обширных исследованиях, доказал тожественность растительных и животных К., чем сильно подвинул изучение животной эмбриологии и гистологии, которая до него развивалась несравненно медленнее, чем растительная. Наконец, последний успех в истории познания К. и вообще построения организмов заключается в открытии сложных процессов деления ядра. Процессы эти, морфологически хорошо изученные с конца семидесятых годов, с физиологической точки зрения еще вовсе не выяснены.
Литература. "Основание к изучению микроскопической анатомии человека и животных", под редакцией М. О. Лавдовского и Ф. В. Овсяникова (2 т., СПб., 1887—1888); "Клетки и ткани. Основы общей анатомии и физиологии", Оскара Гертвига, перевод с немецкого (т. I, СПб., 1894). В указанных сочинениях находится обширный литературный список.
А. Бекетов.
|
Page was updated:Tuesday, 11-Sep-2012 18:15:34 MSK |