| [ начало ] | [ М ] |
Млекопитающие*
(Mammalia) — высший класс позвоночных животных. Основные их признаки следующие: тело покрыто волосами; обе пары конечностей служат по большей части как ноги; череп сочленен с позвоночником двумя затылочными бугорками; нижняя челюсть сочленяется непосредственно с черепом; сердце, как и у птиц, представляет полное разделение артериального и венозного отделов; за двумя исключениями (утконоса и ехидны), все живородящи и вскармливают детенышей молоком.
I. Скелет. Позвоночник М. более резко разграничен на отделы, чем у птиц и пресмыкающихся; он состоит из отделов шейного, грудного, поясничного, крестцового и хвостового. Только у китообразных и сирен (Sirenia), лишенных задних конечностей и тазового пояса, не существует и крестца, и поясничный отдел непосредственно переходит в хвост. Шейный отдел состоит всегда из семи позвонков, независимо от длины шеи, будет ли это шея жирафы или медведя; исключений очень немного: у ламантина (Ma n atus) и ленивца (Choloepus Hoffmanni) обыкновенно лишь шесть шейных позвонков, у ленивцев из рода Bradypus до 9. Первый шейный позвонок (atlas) лишен остистого отростка и снабжен широкими крылообразными поперечными отростками, при основании которых спереди лежат две суставные ямки для двух затылочных бугорков черепа. В сочленении черепа с атласом происходит лишь опускание и поднимание головы; поворот головы направо и налево происходит в сочленении атласа со вторым шейным позвонком (epistropheus), причем атлас вместе с черепом вращается, как вокруг оси, вокруг зубовидного отростка второго позвонка. Зубовидный отросток (processus odontoideus) морфологически соответствует телу атласа, но вполне срастается с телом второго позвонка; на передней стороне его находится гладкая суставная поверхность для сочленения с передней дужкой атласа. В этом сочленении зубовидный отросток укреплен с помощью волокнистой поперечной связки, отделяющей его от канала, заключающего спинной мозг. Грудные позвонки обыкновенно обладают сильно развитыми остистыми отростками, особенно у животных, имеющих длинную шею, служащими для прикрепления затылочной связки (ligamentum nuchae), которая своим передним концом прикрепляется к остистому отростку второго шейного позвонка и к гребню затылочной кости и поддерживает тяжелую голову. Грудных позвонков, к которым прикреплены ребра, бывает обыкновенно 12—15, но может быть и меньше (у некоторых броненосцев всего 9) и больше (у лошади 18, у носорога 19—20, у некоторых ленивцев даже 24). Поясничных позвонков бывает от 8 до 5 и в некоторых случаях даже до 2. Крестец у многих сумчатых состоит всего из двух позвонков; в большинстве же случаев он состоит из 3 или 4, реже 5—6 тесно сросшихся позвонков, из которых обыкновенно лишь первый и частью второй соединены с подвздошными костями. Наиболее значительным колебаниям подвержено у М. число хвостовых позвонков, которых может быть от 4 (у человека и некоторых обезьян) до 46 (у ящера, Manis macrura). Последние хвостовые позвонки не имеют ни верхних дуг, ни отростков и состоят лишь из одних тел, напоминающих суставы пальцев. Ребра М. на верхнем (спинном) конце представляют двойное соединение с позвоночным столбом: головки их сочленяются с суставной ямкой, образованной обыкновенно двумя соседними телами позвонков, а бугорки сочленены с поперечными отростками заднего из обоих позвонков. Нижние концы ребер у большинства М. в течение всей жизни остаются хрящевыми; в тех случаях, когда ребра вполне окостеневают, напр. у китообразных, у неполнозубых, грудинные отделы их являются в виде самостоятельных костей. Только передние, так назыв. истинные, ребра прикреплены к грудной кости; задние, так назыв. ложные, ребра или соединяются с грудинными концами передних, или оканчиваются свободно в брюшной стенке тела. У китообразных последние ребра не соединены даже с позвоночным столбом. Истинных ребер бывает обыкновенно больше, чем ложных. У некоторых М., особенно у неполнозубых, ребра бывают очень широки; у одного муравьеда (Myrmecophaga didactyla) они даже надвигаются своим задним краем на передний край следующего ребра, так что почти не остается межреберных промежутков.
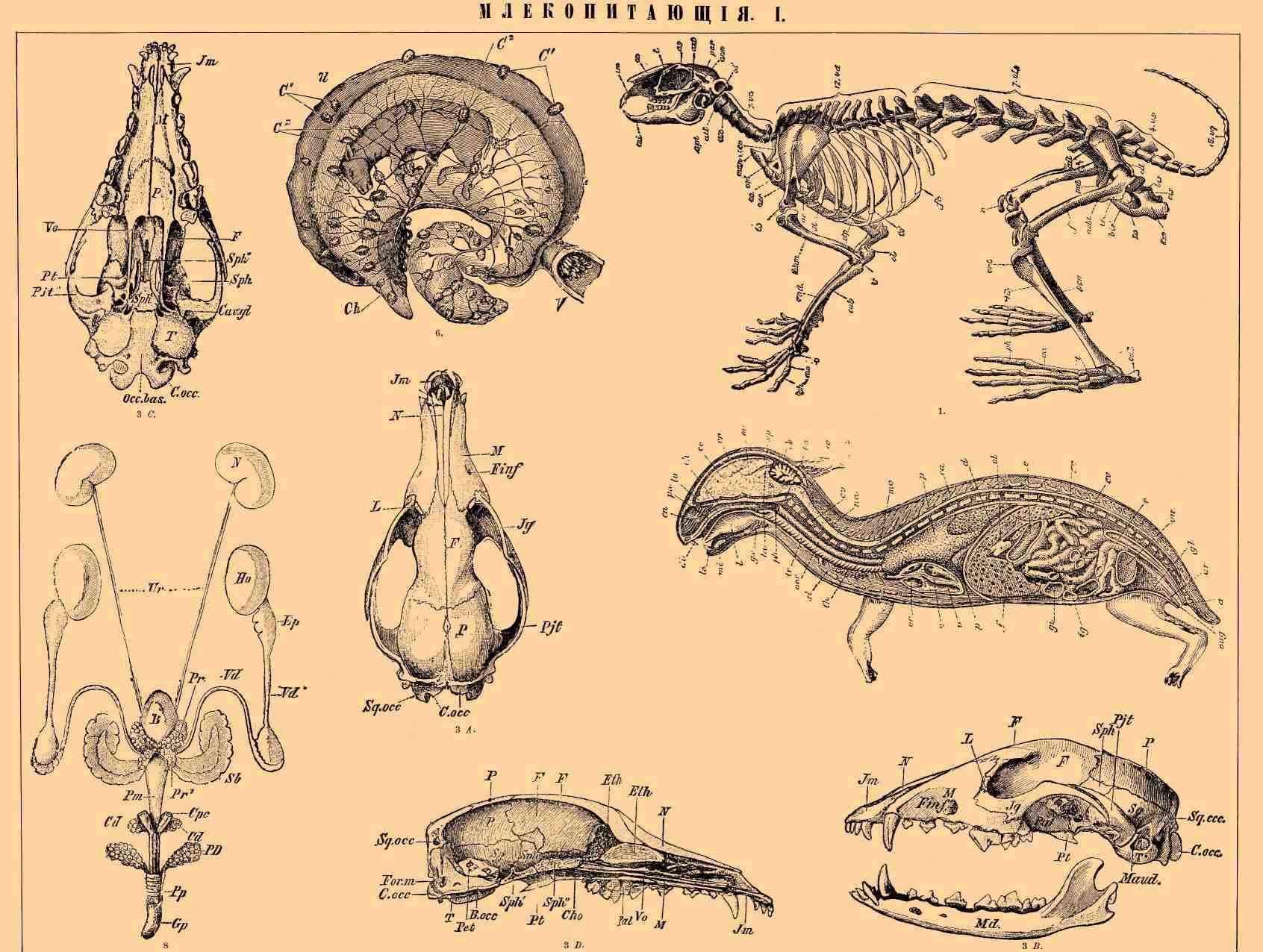
МЛЕКОПИТАЮЩИЕ. I. 1) Скелет кролика: vc — шейные позвонки, atl — атлас, ах — второй шейный позвонок, vd — спинные позвонки, vl — поясничные позвонки, vs — крестцовые позвонки, vq — хвостовые позвонки, со и со' — истинные ребра, fс — ложные ребра, st — грудная кость, man — рукоятка грудной кости, хiр — мечевидный отросток, os — затылочная кость, par — теменные кости, as — бровная дуга, on — носовые кости, im — межчелюстная кость, mi — нижнечелюстная кость, apt — крыловидный отросток, аст — суставной отросток, azt — скуловой отросток височной кости, l — слезная кость, от — лопатка, to — головка лопатки, eо — гребень лопатки, acr — акромион, ас — крючковатый отросток, hum — плечевая кость, tr — блоковидный суставной конец ее, ol — локтевой отросток, cub — локтевая кость, rad — лучевая кость, с — запястье, тc — пясть, ph — суставы пальцев, сil — тело подвздошной кости, ail — крыловидная часть ее, pub — лонное соединение, tis — бугор седалищной кости, bis и bis'— верхняя и нижняя ветвь седалищной кости, асbt—вертлужная впадина, to — foramen obturatum, f — бедренная кость, tr — ее наружный мыщелок, r — коленная чашечка, tib — большая берцовая кость, per — малая берцовая, t — предплюсна или пятка, cal — пяточная кость, mt — плюсна, ph — суставы пальцев.
2) Продольный разрез новорожденного кролика, показывающий расположение внутренностей: di — резцы, le — губы, mi — нижняя челюсть, l — язык, ра — небо, ch — хоаны, cn — носовая полость, cr — свод черепа, bo — основная часть затылочной кости, so — верхняя часть затылочной кости, sp — основная кость, cv — тела позвонков, па — дуги позвонков, еа — остистые отростки, st — грудная кость, lо — обонятельные доли, се — полушария большого мозга, mc — средний мозг, cb — мозжечок, та — продолговатый мозг, те — спинной мозг, la — гортань, tr — дыхательное горло, рр — лопасти легкого, th — грудная железа (thymus), ph — глотка, gs — слюнная железа, oes — пищевод, е — желудок, ig — тонкая кишка, gi — толстая кишка, r — прямая кишка, а — заднепроходное отверстие, f — печень, vb — желчный пузырь, d — грудобрюшная преграда, с — сердце, or — предсердие, v — желудочки, ао — брюшная аорта, vu — мочевой пузырь, иr — мочеиспускательный канал, gl — коуперовы железы, оиg — мочеполовое отверстие.
3) Череп собаки: А — сверху, В — сбоку, С — снизу, D — продольный разрез, Jm — межчелюстная кость, N — носовая кость, М — верхнечелюстная кость с подглазничным отверстием (Foramen infraorbitale — Finf.), Jg — скуловая кость, Pjt — скуловой отросток височной кости, L — слезная с слезным каналом, P — теменная (Р на рис. С. — небная), Sq. осc — чешуя затылочной кости, С оcc — затылочные мыщелки, В. осc — основная затылочная, Pal — небная, Pt — крыловидная, Sph — крылья основной кости, Sph' — основная кость, Sph" — пресфеноидная, Sq — чешуя височной кости; Maud — наружный слуховой проход, T — барабанная, For. т — затылочная дыра, Pet — каменистая кость, Cho — хоаны, Vo — сошник, Eth — вертикальная пластинка решетчатой кости, Eth' — решетчатая пластинка ее, Cav gl — сочленовная ямка для нижней челюсти.
6) Матка коровы с зародышем: V — влагалище, U — матка, Ch — хорион, C' — маточные, С 2 — зародышевые дольки.
8) Мочеполовой аппарат самца ежа: N — почка, Ur — мочеточники, В — мочевой пузырь, Hо — семенные железы, Ер — придаточная часть их, Vd' и Vd — выводной проток, Рr и Рr' — предстательная железа, Sb — семенные пузырьки, Pm — перепончатая часть мочеиспускательного канала, Срс — пещеристые тела, Cd — кауперовы железы, PD — железы крайней плоти, Рр — крайняя плоть, Gp — головка полового члена.
Грудная кость, состоящая обыкновенно из нескольких косточек, у большинства М. узкая и длинная; у летучих мышей и у роющихся в земле животных (кроты, броненосцы) на передней поверхности грудной кости выступает продольный гребень, служащий, как у птиц, для прикрепления грудных мышц.
Кости черепа, М. по большей части разделены швами, лишь в позднем возрасте иногда зарастающими. Только у однопроходных кости черепа сливаются между собой, как у птиц. Число отдельных костей черепа меньше, чем у низших позвоночных; о строении и составе черепа см. Череп.
Плечевой пояс состоит из двух лопаток и двух ключиц. Лопатка представляет собою обыкновенно плоскую треугольную кость, несущую на наружной стороне продольный гребень, который оканчивается спереди над суставной ямкой особым отростком (acromion). Ключицы у многих М. (напр. у копытных) вовсе нет, у других (большинство хищных, некоторые грызуны) они рудиментарные и могут заменяться связками; у тех же М., передние конечности которых способны к разнообразным движениям, ключицы хорошо развиты и соединяют акромиальный отросток лопатки с верхушкой грудной кости. Так назыв. вороньи, или коракоидные, кости, которые у других позвоночных соединяют лопатку с грудной костью, между М. существуют только у однопроходных (Monotremata). У остальных М. зачаток коракоидной кости, появляющийся при эмбриональном развитии, срастается с лопаткой в одну кость, образуя ее отросток, отходящий от нижне-переднего края суставной ямки. Таз М. состоит из трех костей (подвздошной, лобковой и седалищной), срастающихся в одну безымянную кость. Подвздошные кости соединены с позвонками крестца; на брюшной стороне обе половины таза соединены между собой по срединной линии посредством хрящевой или костной спайки, причем у одних млекопитающих, особенно у высших отрядов, сращение ограничивается одними лобковыми костями, у других простирается и на седалищные кости; в некоторых же случаях обе половины таза соединены посредством связок. У сирен и китообразных, лишенных задних конечностей, существуют лишь незначительные рудименты таза в виде двух небольших продолговатых косточек, не соединенных ни между собой, ни с позвоночным столбом. У сумчатых и однопроходных существуют еще особые так назыв. сумочные кости (ossa marsupialia), пара длинных плоских костей в брюшной стенке тела, которые задним концом соединены подвижно с передним краем лобковых костей, а впереди расходятся. Передние конечности М. служат по большей части, как и задние, для движения по земле; но они могут также быть превращены в ласты (у тюленей), в плавники (у сирен и китообразных), быть приспособлены к хватанию (обезьяны и человек) или к летанию (летучие мыши). Плечевая кость у всех плавающих М. сильно укорочена; она коротка также у копытных, у которых зато сильно удлинена кисть передней конечности. У роющих землю М. плечевая кость широкая и с сильно развитыми отростками. Из двух костей предплечья локтевая (ulna) служит преимущественно для сочленения предплечья с плечом; лучевая (radius), напротив, принимает главнейшее участие в сочленении кисти с предплечьем. У многих низших М., напр. у некоторых грызунов и хищных, обе кости предплечья, лучевая и локтевая, развиты почти одинаково и соединены между собою свободно, так что нижний конец лучевой кости может несколько перемещаться около локтевой. У обезьян и человека лучевая кость, которая почти одна участвует в сочленении с кистью, достигает наибольшей подвижности и дает возможность вращательным движениям кисти руки. Лучевая кость вращается при этом вокруг продольной оси, проходящей через ее верхний конец и через нижний конец локтевой; из своего первоначального положения, рядом с локтевой, на наружной стороне предплечья, лучевая кость становится накрест с локтевой, переходя своим нижним концом на внутреннюю сторону предплечья; кисть руки, прикрепленная к лучевой кости, также вращается при этом вокруг своей оси: при параллельном положении костей предплечья кисть руки смотрит ладонью вверх (supinatio); когда лучевая кость перекрещивается с локтевой, кисть руки поворачивается ладонью вниз (pronatio). У тех М., у которых передние ноги служат исключительно для опоры тела, а таких большинство, — кисть передней конечности не способна к вращательным движениям и укреплена постоянно ладонью вниз. При этом обе кости предплечья соединены между собою неподвижно, причем лучевая кость развивается в ущерб локтевой, которая оттесняется на заднюю сторону предплечья, срастается с лучевой, становится нередко недоразвитой (у жвачных); у верблюдов и у однокопытных от локтевой кости остается только ее верхний отдел, слитый с лучевой костью и несущий сильный локтевой отросток (olecranon). Кисть передней конечности состоит из запястья, или предпястья (carpus), пясти (metacarpus) и пальцев, различным образом развитых. Запястье состоит всего из семи костей, расположенных в два ряда, по три и четыре; очень часто некоторые из этих косточек между собою сливаются. У некоторых М., у грызунов, насекомоядных, полуобезьян, у орангутанга между первым и вторым рядом костей запястья лежит еще особая центральная кость (carpale centrale); она появляется также у зародыша человека, но при дальнейшем развитии сливается с ладьевидной костью запястья (radiale).
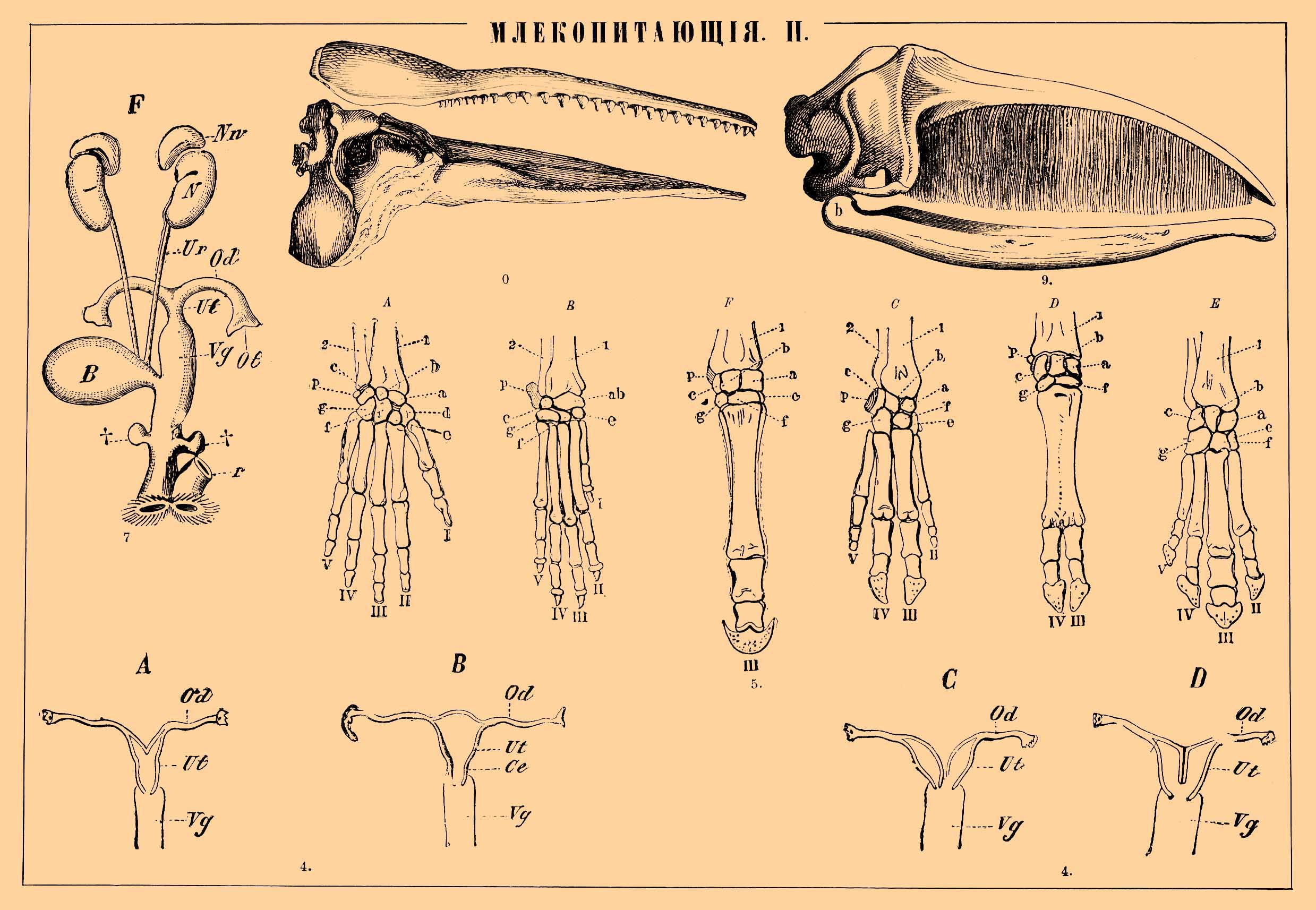
4) Различные формы матки: А — двурогая, В — простая, С — двойная матка, D — двураздельная.
5) Скелет кисти различных млекопитающих: А — человека, В — собаки, С — свиньи, D — быка, E — тапира, F — лошади. 1 — нижний конец лучевой кости; 2 — нижний конец локтевой; a, b и с — три кости первого ряда запястья: а — ладьевидная, b — полулунная, с — трехгранная, ab — результат слияния а с b; p — гороховидная кость; d, е, f и g — четыре кости второго ряда запястья: d — большая многогранная, е — малая многогранная, f — головчатая, g — крючковатая; I—V — пять пальцев.
7) Мочеполовой аппарат самки ежа: Nn — надпочечные железы, N — почки, Ur — мочеточник, В — мочевой пузырь, Od — яйцевод, Ot — отверстие яйцевода, Ut — матка, Vg — влагалище, †† — придаточные железы, r — прямая кишка.
9) Череп гренландского кита (Balaena mysticetus) с пластинками "китового уса" сбоку.
10) Череп кашалота (Physeter macrocephalus) сбоку.
Длинные кости пясти — у пятипалых пять, с уменьшением числа пальцев уменьшается и их число — сочленяются на одном конце с костями второго ряда запястья, на другом с пальцами. Пальцев у М. бывает не больше пяти и обыкновенно менее. Из них первый, считая с внутренней или лучевой стороны, или большой, состоит из двух, остальные из трех суставов, или фаланг; только у китообразных число суставов на втором и третьем пальцах увеличено. При уменьшении числа пальцев чаще всего недостает большого пальца. У парнопалых преобладающее развитие получают два средних пальца, третий и четвертый, а крайние пальцы — второй и пятый — недоразвиты или исчезают. У непарнопалых преобладающее развитие перед всеми остальными получает третий палец, и в крайнем случае — у однокопытных — только он один и остается. Параллельно с сокращением числа пальцев уменьшается и число костей пястья и запястья. У жвачных преобладающее развитие получают два пальца, 3-й и 4-й; соответствующие им кости пясти сливаются в одну длинную кость, а боковые (2-я и 5-я) пястные кости делаются зачаточными или даже совсем исчезают. У однокопытных единственному развитому пальцу соответствует и одна кость пясти, по бокам которой сохраняются в виде маленьких, так назыв. грифельных, косточек рудименты верхнего отдела второй и четвертой косточек пясти. В задних конечностях бедренная кость, которая у человека является самой длинной из костей скелета, у многих М. бывает довольно коротка и уступает голени в длине. Из двух костей голени преобладающего развития достигает обыкновенно большая берцовая (tibia), между тем как малая берцовая (fibula) нередко, особенно у копытных, становится рудиментарной. У жвачных от малоберцовой остается лишь нижний конец, сочлененный с большеберцовыми костями и с предплюсною. У лошади остаток малоберцовой кости в виде короткого, тонкого костяного столбика находится на верхнем конце большой берцовой. У многих грызунов и насекомоядных обе кости голени на большем или меньшем протяжении срастаются между собою. У некоторых лазящих сумчатых, у которых на задних ногах большой палец может противополагаться остальным, большая берцовая может до известной степени вращаться около малой берцовой, наподобие вышеописанного вращения лучевой ок. локтевой. Надколенная чашечка, прикрывающая спереди коленный сустав и лежащая в сухожилии большого разгибателя голени, отсутствует только у некоторых сумчатых. Предплюсна (tarsus) в полном развитии состоит, как и запястье, из семи костей. Первый ряд костей предплюсны составляют две кости: таранная (astragalus), которая по большей части одна участвует в сочленении стопы с голенью, и пяточная (calcaneus), снабженная более или менее сильно выдающимся назад отростком, так назыв. пяточным бугром, к которому прикрепляется Ахиллесово сухожилие. За первым рядом костей предплюсны следует ладьевидная кость (naviculare), соответствующая центральной кости запястья, и затем второй ряд костей предплюсны, числом четыре, сочлененных с костями плюсны. Иногда некоторые кости предплюсны сливаются между собою. Кости плюсны (metatarsus) и пальцы задних конечностей бывают обыкновенно в том же числе и повторяют характер передних конечностей; часто, однако, на задних ногах меньше пальцев, чем на передних. Так, в семействах собак и кошек на передних ногах пять пальцев, на задних четыре. У многих сумчатых, у полуобезьян и обезьян большой палец задних ног может противопоставляться остальным, а в таком случае и задние конечности могут служить для хватания. Одни М. при передвижении касаются земли всей стопою задней, всею кистью передней ноги; другие — лишь фалангами пальцев, тогда как кости плюсны и пясти приподняты над землею; первые назыв. стопоходящими (напр. медведь, еж), вторые пальцеходящими (собака, кошка). Копытные М. опираются на землю лишь верхушками последних суставов пальцев.
II. Волосы и другие производные кожи. Кожные железы. Молочные железы. Присутствие волос на теле составляет такой же характерный признак для класса М., как перья для птиц (см. Волосы). Лишь немногие толстокожие М., как слоны, носороги, бегемоты, почти лишены волос; у китообразных лишь на верхней губе есть немного волос, и то часто только у зародышей. Кроме мягких и гибких волос, могут быть жесткие и тугие щетины; иногда волосы могут быть превращены в иглы (еж, дикобраз). У одних М. (напр. быки, лошади) тело покрыто одинаковыми волосами (шерстью), достигающими лишь особенной длины в хвосте и в гриве; у других (напр. куница, кошка) мех состоит из двух различных сортов волос: из густых, мягких, тонких и коротких волос, составляющих пух, или подшерсток, и из более длинных и толстых и более редких волос, покрывающих первые и составляющих собственно шерсть, или ость; часто они и по цвету отличаются от пуховых волос. У ящера (Manis) из отряда неполнозубых спина и бока покрыты большими роговыми чешуями, прикрывающими друг друга задними концами наподобие черепицы: чешуи эти могут подыматься как иглы; между ними сидят жесткие волосы, и все брюхо покрыто волосами. Чешуи эти — особое видоизменение волос, представляющие как бы сросшиеся рядами иглы. У многих грызунов (бобр, крыса), сумчатых и насекомоядных на хвосте мелкие роговые чешуйки, похожие на роговые чешуйки пресмыкающихся. Подобное же сходство с соответственными образованиями пресмыкающихся представляют роговые выросты кожи на концах пальцев — ногти, когти и копыта М. У разных М. существуют еще особенные роговые образования кожи: полые рога антилоп, быков и овец, сидящие на особых выростах лобных костей; рог носорога, образованный из многих склеенных между собою волокон; небные пластинки (так назыв. ус) китов и друг. Костяные образования в коже находятся только у броненосцев, в виде щитков, расположенных поперечными рядами, образующих костяной панцирь, покрывающий большую часть тела и снаружи прикрытый роговым слоем. Из костной ткани, образующейся в коже, состоят также рога оленей, периодически отпадающие и возобновляющиеся. В коже М. находятся двоякого рода железы: потовые и сальные. Первые состоят из цилиндрических трубочек, нижний конец которых лежит в соединительнотканном слое кожи и свернут здесь в клубочек, а прямой выводной канал проникает в эпидерму и открывается наружу. Сальные железы, мешковидной или гроздевидной формы, открываются обыкновенно в сумку волоса, часто по нескольку в одну. Оба рода желез у разных М. и на разных частях тела представляют различные видоизменения; часто они выделяют особые пахучие вещества, играющие важную роль в жизни животного (мускусные железы). Молочные железы также следует отнести к органам кожи; каждая молочная железа состоит из скученной массы отдельных кожных желез, которые и по своему строению, и по способу образования своего отделения сходны с сальными железами. Лишь у однопроходных (утконоса и ехидны) молочные железы построены по типу трубчатых желез и, следов., сходны с потовыми железами, а не с сальными. У ехидны молочные железы открываются в два небольших углубления в боковых сторонах мешка, служащего для вынашивания детеныша (молочные карманы). Подобные же кармашки — углубления кожи — образуются при начале развития молочных желез у зародышей высших М.; затем на их место развиваются сосцы. Однако у некоторых M. (Antilope cervicapra, в известной степени и мыши) молочные карманы сохраняются и во взрослом состоянии. Число сосцов и положение их у различных М. весьма изменчиво; их может быть от одной до семи пар, они могут лежать в паховой области (копытные, китообразные), или тянуться двумя рядами вдоль груди и брюха (хищные, свиньи), или находиться исключительно на груди (слоны, сирены, многие полуобезьяны, летучие мыши и приматы).
Грудобрюшная преграда, или диафрагма, составляет один из характерных анатомических признаков М. Это сводообразная мускулистая перегородка, отделяющая грудную полость от брюшной. Она прикреплена к позвоночному столбу, ребрам и грудной кости; в вершине ее выпуклой части лежит центральная сухожильная пластинка. Диафрагма играет большую роль в процессе дыхания.
III. Нервная система. Органы чувств. Головной мозг М. по сравнению с птицами и пресмыкающимися отличается особенным развитием переднего мозга и мозжечка сравнительно с другими отделами. При этом между полушариями большого мозга и мозжечка развивается система сильных поперечных комиссур (мозолистое тело — corpus callosum, свод — fornix, Варолиев мост), существующая у других позвоночных лишь в зачатке. У низших М. (однопроходных, сумчатых, насекомоядных, неполнозубых, грызунов и рукокрылых) полушария большого мозга еще не закрывают сверху среднего мозга (четыреххолмия); у хищных и копытных большой мозг достигает назади до мозжечка, а у обезьян и у человека прикрывает сверху мозжечок. Поверхность полушарий большого мозга остается у утконоса, у многих сумчатых и неполнозубых совершенно гладкою, как у других позвоночных; у остальных М. на поверхности полушарий появляются выпуклые извилины (gyri), разделенные бороздами, достигающие наибольшего развития у обезьян и у человека. Спинной мозг М. обыкновенно оканчивается в крестцовой или поясничной области, иногда даже (у ежа, ехидны, рукокрылых) в грудной области позвоночника; нервы, выходящие из самой задней части спинного мозга, тянутся друг подле друга в позвоночном канале, образуя так наз. конский хвост. В местах выхода нервов для конечностей спинной мозг представляет утолщения: шейное и поясничное; последнее особенно развито у М. с сильно развитыми задними конечностями (кенгуру) и продолжает существовать у китообразных, лишенных задних конечностей. Органы чувств. Главным органом осязания М. служат губы с длинными, тугими осязательными волосами, сумки которых богаты нервами; органом осязания служит также верхушка носа, удлиненная у некоторых зверей в хобот. У обезьян и человека осязание особенно развито также в концевых подушечках пальцев; значительная подвижность пальцев и способность противопоставления большого пальца остальным делает у них, особенно у человека, из передней конечности руку, специальный орган для схватывания предметов и для ощупывания. Орган обоняния достигает у М. высшего развития между позвоночными. Значительное развитие лицевых костей придает носовой полости большую емкость; вертикальной перегородкой она делится на две половины, открывающиеся наружу спереди (ноздри) и в глотку сзади (choanae). Боковые стенки носовой области (решетчатая кость) образуют множество небольших ячеек (лабиринт) и разнообразные выступы внутрь носовой полости, так наз. раковины. Но только верхний отдел носовой полости, соответствующий верхней и средней раковине и верхней части вертикальной носовой перегородки, является органом обоняния — именно заключает в своей слизистой оболочке разветвления обонятельного нерва. Нижний же отдел носовой полости с нижней раковиной является лишь дыхательною частью носовой полости, выстлан обыкновенным мерцательным эпителием и иннервируется тройничным нервом. Впереди носовых костей, в стенках переднего отдела носовой полости, лежат хрящевые пластинки, или самостоятельные (у жвачных, у лошадей), или отходящие от хрящевой перегородки носа (у хищных); хрящи эти вместе с носовыми костями обусловливают внешнюю форму носа. Наружные носовые отверстия могут расширяться и суживаться помощью особых мускулов (их нет только у утконоса); у тюленей они могут даже вполне закрываться. У некоторых М. (свиньи, тапиры, слоны, кроты, землеройки, выхухоль) наружный нос удлиняется в хобот, достигающий особенных размеров у слонов, которым он служит органом хватания. У китообразных органы обоняния недоразвиты. О строении органов слуха и зрения см. Глаз и Ухо. Упомянем лишь о наружном ухе М., имеющем волокнисто-хрящевой остов и снабженном мышцами, приводящими его в движение. Остов уха состоит из одной хрящевой пластинки или даже из трех хрящей. Форма и величина уха чрезвычайно различны. Совсем нет наружного уха у однопроходных, у многих М., живущих в воде (сирены, китообразные; большинство ластоногих), у некоторых ведущих подземную жизнь (крот). В наружном ухе человека сравнительно с обезьянами дистальная часть (так наз. ушная складка) его редуцирована, именно край ее завернут внутрь, тогда как основная часть ("пояс ушных бугорков") не обнаруживает признаков регресса в развитии.
IV. Зубы существуют у всех М., кроме однопроходных, части китообразных и родов ящер (Manis) и муравьед (Myrmecophaga) из неполнозубых. У беззубых китообразных при развитии зародыша появляются в челюстях зачатки зубов, но они не развиваются дальше и вскоре исчезают. У утконоса есть зубы лишь в молодости и вскоре выпадают. Зубы М. сидят всегда корнями в ячейках челюстных костей. Различают резцы (dentes incisivi); клыки (d. canini), передние, или ложнокоренные (praemolares), и истинные коренные (molares); зубы одной челюсти приходятся обыкновенно против промежутков между зубами другой. Резцы сидят наверху в межчелюстной кости, внизу по обе стороны места сращения двух половин нижней челюсти и имеют обыкновенно долотообразную форму. Иногда их не бывает вовсе, напр. у жвачных в верхней челюсти. У слонов в нижней челюсти нет резцов, в верхней два громадных резца принимают форму так называемых бивней. Клыков бывает 4, по одному в каждой половине челюсти, конической более или менее изогнутой формы; они развиты особенно у хищных М., у травоядных их часто вовсе не бывает, и тогда между резцами и коренными остается значительный промежуток. Коренные зубы представляют особенно резкие различия по числу и строению. Иногда (у некоторых копытных, у грызунов) все коренные зубы приблизительно одинаковы, чаще же различают передние ложнокоренные зубы от задних истинных коренных. Коренные зубы снабжены обыкновенно крепкой, широкой коронкой с плоской бугорчатой поверхностью. Каждый такой зуб соответствует нескольким простым коническим зубам (каковы зубы пресмыкающихся); при эмбриональном развитии несколько зубных сосочков сливаются для образования одного коренного зуба. У хищных М. коронка коренных зубов острая, режущая, и один или два из средних коренных зубов отличаются своей величиной — так назыв. плотоядный зуб. Для обозначения числа зубов пользуются особой формулой, ставя над чертой число зубов в половине верхней челюсти, под чертой — в половине нижней челюсти. Так, зубам человека и обезьян Старого Света соответствует формула: (2. 1. 2. 3.)/(2. 1. 2. 3.) т. е. резцов 2/2, клыков 1/1, ложнокоренных 2/2, истинно-коренных 3/3 . Общее число зубов, следовательно, будет двойное. Зубная формула лошади 3/3. 1/1. 3/3. 3/3., коровы 0/3. 0/1. 3/3. 3/3. Только у некоторых китообразных и у неполнозубых зубы, первоначально развившиеся у зародыша, остаются на всю жизнь; у остальных М. происходит смена зубов, замена первых, молочных, зубов постоянными. Об этом см. в статье Зубы. Число сменяемых зубов и время их смены различны. У человека и у многих других смене подлежат все резцы, клыки и передние коренные (praemolares). У сумчатых в каждой половине челюсти сменяется только один коренной зуб. Между этими крайними случаями наблюдается целый ряд переходных степеней. В некоторых случаях, напр. у тюленей, смена молочных зубов постоянными происходит вскоре после рождения или даже раньше его, так что молочные зубы совсем не функционируют. У зубастых китообразных, напротив, остаются на всю жизнь молочные зубы, как видно из того, что у них появляются, но не развиваются далее зачатки постоянных зубов.
V. Пищеварительные органы — см. ст. соответ. слов. Здесь упомянем только, что развитие подвижных мускулистых губ делает из них часто (напр. у копытных) органы для захватывания пищи. Полость между губами, щеками и челюстями (преддверие рта, vestibulum oris) у многих узконосых обезьян, грызунов и сумчатых образует боковые мешковидные выступы, защечные мешки. Язык, отличающийся сильно развитою мускулатурою, служит не только органом осязания и вкуса, но также и для проглатывания пищи. Обыкновенно он имеет плоскую, лентообразную форму и может высовываться изо рта; особенно длинный и узкий язык у ехидны и у муравьеда. Поверхность языка может быть или гладкая, или покрытая мягкими сосочками, или (у многих хищных, напр. у кошачьих) усаженная направленными назад роговыми шипами. Желудок имеет вид простого овального или продолговатого мешка, лежащего более или менее наискось; он может быть разделен на два или более отделов. Особенного развития это разделение достигает у жвачных (см.).
VI. Органами дыхания М. служат легкие. Гортань М. сравнительно с другими позвоночными отличается присутствием двух особых хрящей: щитовидного и надгортанного. У всех М., кроме китообразных, в полости гортани находятся две складки слизистой оболочки, так наз. голосовые связки, ограничивающие голосовую щель. У большинства существует еще вторая, вышележащая пара таких складок, более слабых, так наз. верхние или ложные голосовые связки. Между верхними и нижними складками находятся мешковидные углубления, так наз. Морганьевы мешки или желудочки. Иногда, напр. у некоторых обезьян, Морганьевы мешки достигают значительного объема, вытягиваются из гортани и тянутся более или менее далеко вниз по шее; особенно сильного развития достигают Морганьевы мешки у американского ревуна (Mycetes). Дыхательное горло делится на две главные ветви, бронхи, к которым присоединяется еще иногда небольшой дополнительный третий бронх. Легкие (см.) по большей части разделены на лопасти, причем обыкновенно правое легкое больше левого и разделено на большее число лопастей. Каждое легкое помещается в особой замкнутой полости, выстланной особой перепонкой, так наз. грудной плевой или плеврой.
Сердце М., как у птиц, разделено сплошною продольною перегородкою на два отдела, левый и правый, и состоит из двух предсердий и двух желудочков. См. Сердце и Кровообращение. Сердце лежит в особом мешке (околосердечная сумка, pericardium); у человека и у обезьян оно имеет косое положение, отходя влево от срединной плоскости тела, и околосердечная сумка лежит на грудобрюшной преграде, срастаясь с ее сухожильным центром. У других М. положение сердца почти совпадает с срединной: плоскостью тела, и околосердечная сумка часто совсем не касается грудобрюшной преграды.
М. принадлежат (вместе с птицами) к так назыв. теплокровным животным или животным с постоянною температурою тела, которая может колебаться у них лишь в весьма незначительных пределах. Так, нормальная средняя темп. лошади 37,5° Ц., кошки 38,3, собаки около 39. У мелких М. температура тела обыкновенно выше, чем у крупных, и может подлежать большим колебаниям. Замечательное отклонение от прочих М. представляют и в этом отношении однопроходные, отличающиеся сравнительно низкою температурою тела и значительными колебаниями ее (у ехидны от 26 1/2 до 34° Ц.), Некоторые М. (еж, хомяк, сурок, медведь, барсук и др.) проводят зиму в состоянии так назыв. зимней спячки, в течение которой работа сердца и дыхание падают до самой незначительной величины и температура тела может понизиться до нескольких градусов. Почки М. лежат в поясничной области, по бокам позвоночника; правая почка бывает нередко выше левой. Иногда (китообразные, ластоногие, выдра, медведь и др.) почки имеют гроздевидный характер и состоят из отдельных лопастей, соединенных лишь ветвями мочеточника; в других случаях (гиены, кошки, бык, слон) лопасти почек срастаются между собою, но остаются заметными на поверхности; наконец, в третьих поверхность почек совершенно гладкая. Мочеточники, выходя из почек, тянутся на довольно значительном протяжении через брюшную полость и впадают в мочевой пузырь; только у однопроходных мочеточники независимо от мочевого пузыря впадают в мочеполовой синус. Последний открывается у однопроходных вместе с прямой кишкой в общую клоаку — чем эти животные резко отличаются от всех остальных М., у которых заднее отверстие кишечника всегда обособлено от мочеполового отверстия. Однопроходные приближаются в этом отношении к низшим позвоночным, пресмыкающимся и птицам.
VII. Половые органы. Семенные железы (testiculi) самцов в эмбриональный период лежат в брюшной полости около почек. В иных случаях они и остаются на всю жизнь в брюшной полости, или на своем первоначальном месте (однопроходные), или несколько позади почек (китообразные, некоторые неполнозубые, слоны и др.). У других М. первоначальное положение семенников постепенно изменяется, они опускаются вниз и лежат или в паховой области на брюшной стенке (мозолистоногие, многие грызуны и хищные), или в особом мешке — мошонке. У многих М. семенники могут периодически (по окончании течки) втягиваться из мошонки обратно в брюшную полость. Яичники также парные у всех М., и обыкновенно оба одинаково развиты; лишь у однопроходных, подобно тому как у птиц, правый яичник недоразвит. Форма их приблизительно овальная; поверхность бугорчатая вследствие выпячивающихся Граафовых пузырьков (см.); изредка яичники имеют гроздевидную форму (однопроходные, некоторые грызуны и сумчатые). Яйцевые клетки М., лежащие в названных Граафовых пузырьках, были открыты в 1827 г. Бэром; они очень небольшой величины (около 0,2 мм в диаметре), состоят из зернистой протоплазмы с большим ядром и окружены довольно толстой светлой оболочкой, которая пронизана многочисленными порами. При достижении зрелости Граафов пузырек, в котором лежит яйцевая клетка, лопается, и последняя переходит в брюшную полость, оттуда в яйцевод. Выводные протоки женских половых органов М., развивающиеся из так назыв. Мюллеровых каналов зародыша, состоят из яйцеводов, матки и влагалища. Узкие яйцеводы (фаллопиевы трубы) начинаются широкой воронкой, снабженной по краям зазубренными выростами. У многих М. яйцеводы теснее сообщаются с яичниками; в иных случаях (кенгуру, некоторые грызуны) яичники лежат в самых воронках яйцеводов, в других (летучие мыши, многие хищные) они лежат в особых карманах брюшины, куда открываются воронки яйцеводов. У однопроходных половые протоки (Мюллеровы каналы зародыша) обеих сторон остаются отделенными друг от друга на всем своем протяжении; у сумчатых, они хотя и срастаются в своем нижнем отделе, но их полости остаются раздельными, так что у них находятся две матки и даже два отдельных влагалища. У всех остальных М. существует только одно влагалище благодаря полному слиянию между собою в нижнем конце обоих половых протоков; что касается матки, то она в различной степени сохраняет следы своего происхождения из парных Мюллеровых каналов. У многих грызунов (зайцы; белки) и летучих мышей существуют еще две отдельных матки (двойная матка), открывающихся каждая отдельно во влагалище; у других грызунов обе матки в нижнем конце сливаются и сообщаются с влагалищам одним общим отверстием, но все-таки разделены друг от друга на большей части длины (двухраздельная матка). У китообразных, копытных, насекомоядных и хищных двойственное происхождение матки выказывается лишь еще в том, что она вытянута на переднем конце в два рожка, переходящие в яйцеводы (двурогая матка). У летучих мышей, обезьян и человека матка простая, резко обособленная от обоих яйцеводов и не несущая уже следов своего парного происхождения. Все М., кроме однопроходных, приносят живых детенышей. Лишь однопроходные, утконос и ехидна размножаются яйцами. Их яйца представляют по сравнению с яйцевой клеткой остальных М. громадную величину, достигая до 15 мм длины, и заключены в перепончатую (кератиновую) скорлупу, в которой у утконоса есть даже примесь извести. Утконос, у которого не бывает мешка на брюхе, сносит обыкновенно два яйца, в которых уже сильно развитые зародыши, в подземную нору; ехидна носит с собою единственное снесенное яйцо в особом мешке на брюхе, в котором открываются молочные железы. У всех остальных М. развитие зародышей протекает в матке, при чем наблюдается постепенное усовершенствование питания зародыша на счет организма матери. Дробление яйца и образование зародышевых листов у М. довольно сильно отличаются от этих процессов у пресмыкающихся и птиц благодаря отсутствию питательного желтка в яйце М., который в значительном количестве существует только у однопроходных и сумчатых. Однако развитие яйца показывает, что отсутствие в нем питательного желтка есть явление вторичное, вызванное приспособлением зародыша к питанию на счет тела матери, и что предки современных М. должны были обладать яйцами, подобными яйцам птиц и пресмыкающихся, т. е. богатыми питательным желтком. На это указывает образование в зародышах М. желточного мешка с системой кровеносных сосудов для всасывания желтка, как у птиц, при отсутствии самого желтка. У сумчатых (за одним только известным исключением) зародыш, развиваясь в матке, не соединен с ее стенкой; питание зародыша совершается на счет выделений стенок матки, причем для их поглощения служит большой желточный мешок зародыша. Однопроходных и сумчатых М. противополагали поэтому всем остальным М. под названием беспоследных М. (Apliacentalia). Лишь в самое последнее время у одного сумчатого (Perameles obesula) найдено тесное соединение плода со стенкой матки, причем в образовании плаценты принимает участие, как у Placentalia, мочевой мешок (Aliautois). У всех остальных последных M. (Placentalia) зародыш вступает в тесную связь с слизистой оболочкой матки при помощи выростов наружной, серозной или ворсинчатой, зародышевой оболочки. В эти выросты или ворсинки врастают кровеносные сосуды мочевого мешка (allantois), и когда выросты проникают в стенку матки, то является возможность для зародыша поглощения питательных веществ из крови матери. У одних М. ворсинки имеют простую форму и равномерно распределены по поверхности ворсинчатой оболочки, образуя так наз. разлитую плаценту (послед, детское место, placenta diffusa) — у неполнозубых, китообразных, сирен, непарнокопытных, свиней, бегемотов, верблюдов. У большинства жвачных плацента дольчатая (pl. cotyledonea); длинные и сильно разветвленные ворсинки сидят на ворсинчатой оболочке пучками, образуя так наз. дольки (cotyledones), которых может быть от 5—6 до 1 0—100.
Против каждой дольки слизистая оболочка матки образует утолщение, богатое кровеносными сосудами и пронизанное многочисленными глубокими и разветвленными углублениями, в которые и входят ворсинки. Таким образом, в каждой дольке различается зародышевая часть плаценты, образовавшаяся из выростов оболочки зародыша, от материнской части плаценты, состоящей из измененной слизистой оболочки матки. В углублениях материнской части плаценты стенками матки выделяется особая питательная жидкость, похожая по составу на молоко, так наз. маточное молоко, которая и поглощается ворсинками зародыша. При разлитой и при дольчатой плаценте связь зародыша с маткой слабая; ворсинки могут быть без труда извлечены из ямок слизистой оболочки, в которые они погружены. У всех остальных М. зародышевая и материнская часть плаценты так тесно срастаются, что не могут быть отделены друг от друга. Тогда при рождении детеныша вместе с зародышевыми оболочками отделяется более или менее значительная часть слизистой оболочки матки, образующая отпадающую оболочку (decidua). У М., обладающих отпадающей оболочкой (deciduata), плацента представляет две главные формы: поясная плацента (pl. zonana) и дисковидная (pl. discoidea). При поясной плаценте, существующей у хищных и ластоногих, густые древовидные ворсинки сидят на ворсинчатой оболочке широким кольцом вокруг зародыша. Дисковидная плацента (у насекомоядных, грызунов, рукокрылых, обезьян и человека) имеет вид кружка и занимает лишь небольшую часть поверхности плода, но отличается чрезвычайным развитием ворсинок. При дисковидной и поясной плаценте особенного развития достигают кровеносные сосуды в материнской части плаценты; у обезьян и у человека в ней образуются обширные кровеносные пространства, как бы кровоизлияния в промежутке между стенкою матки и ворсинками зародыша; ворсинки плаценты в этом случае непосредственно обмываются материнского кровью. Продолжительность беременности у М. весьма различна; у слона она длится 20 1/2 месяцев, у лошади 346 дней, у коровы 280, у овцы 150, кролика 30, мыши 21 день. Всего короче беременность у сумчатых, у которых детеныши рождаются недоразвитыми и донашиваются в сумке. Между последовыми М. недолго носят хищные, у которых детеныши рождаются с закрытыми глазами и обыкновенно голыми. Что касается числа единовременно производимых на свет детенышей, то у крупных М., снабженных небольшими числом сосцов, оно невелико — 1, 2. У меньших М., снабженных многими сосцами, оно может, напротив, достигать большого числа: у различных грызунов и у свиней до 10 12, у свиней иногда даже до 20 и более.
VIII. О географическом распространении М. заметим следующее: за исключением некоторых островов Тихого океана, М. водятся по всей земле, но лишь немногие виды космополиты, большинство имеет ограниченную область распространения. Число родов и видов уменьшается при передвижении от экватора к полюсам. Из зоогеографических областей особенностями в фауне М. отличаются неотропическая область, мадагаскарская подобласть и австралийская область. Австралийской области исключительно свойственны однопроходные и сумчатые (с единственным исключением двуутробок, Didelphyidae); для Мадагаскара и смежных островов характерны полуобезьяны, для Южной Америки ей одной свойственный (за исключением двух родов: Manis и Orycteropus) неполнозубые. Древнейшие ископаемые остатки М. попадаются в триасе и юре; они состоят обыкновенно из нижней челюсти и даже из одних зубов и причисляются, хотя часто и без достаточных оснований, к сумчатым. Плацентарные М. появляются лишь с началом третичного периода; особенно богата древнейшая эоценовая фауна в Северной Америке. Значительная часть этих ископаемых М. представляет собой формы, которые не могут быть включены в рамки современных отрядов М., представляя собой соединительные между ними звенья и являясь общими родоначальниками М., относимых в настоящее время к различным отрядам. Так, эоценовые Creodonta являются, по-видимому, общими родоначальниками современных и хищных, и копытных. В миоцене появляются уже все современные отряды М. и большинство нынешних семейств; плиоценовая фауна состоит уже, за немногими исключениями, из современных родов. Современные М. делятся на три подкласса: однопроходные (Monotremata), сумчатые (Marsupialia) и последовые (Placentalia). Последние заключают в себе отряды: неполнозубых (Edentata), китообразных (Cetacea), сирен или травоядных китов (Sirenia), непарнокопытных (Perissodactyla), парнокопытных (Artiodactyla), хоботных (Proboscidea), плоскокопытных (Lamnungia), хищных (Carnivora), ластоногих (Pinnipedia), грызунов (Rodentia), насекомоядных (Insectivora), рукокрылых (Chiroptera), полуобезьян (Prosimiae), обезьян (Primates). К последнему отряду относится и человек [Рисунки по анатомии М., кроме приложенных в этой статье, см. в отдельных статьях по анатомии: головной мозг (фиг. головного мозга человека и кролика — 7 и 8 второй таблицы, а не 8 и 9, как сказано в объяснении), нервная система, сердце и т. д., а также в статьях об отдельных отрядах и меньших группах.]. Ср. Бобрецкий, "Основания зоологии"; Ludwig (Leuniss), "Synopsis d. Thierkunde"; W. H. Flower and R. Lydekker, "An introduction to the study of Mammals living and extinct" (1891); Шимкевич и Полежаев, "Курс зоологии позвоночных"; Брэм, "Жизнь животных"; Wiedersheim, "Vergleich. Anatomie d. Wierbeelthiere" (обширная литература); Гертвиг, "Учебник эмбриологии позвоночных" (обширная литература); Симашко, "Русская фауна"; Bronn, "Klassen u. Ordnungen d. Thierreichs" ("Mammalia" von Giebel und Leche); Vogt und Yung, "Lehrbuch der vergleichenden Anatomie" (II т.).
В. Ф.
|
Page was updated:Tuesday, 11-Sep-2012 18:15:54 MSK |