
| [ начало ] | [ Р ] |
Ракообразные*
(Crustacea). Характеристика и общая организация. — Конечности. — Покровы и мускулатура. — Органы пищеварения. — Нервная система. — Органы чувств. — Кровеносная система. — Органы дыхания. — Выделительные органы. — Половые органы. — Размножение и развитие. — Классификация.
Характеристики и общая организация. Р. класс типа суставчатоногих (см.) или членистоногих (Arthropoda), заключающий в себе громадное число видов, отличающихся разнообразием строения. Р. могут быть кратко охарактеризованы, как водные суставчатоногие, дышащие жабрами и снабженные двумя парами предротовых конечностей, или усиков, на голове, многочисленными ножками на груди, а большей частью и на брюшке. Тело Р. состоит из отдельных сегментов, или члеников, число которых весьма разнообразно и варьирует у низших Р. — Entomostraca, когда как для всех высших P. — Malacostraca — является постоянным. В большинстве случаев эти членики не одинаковы, а разнозначащи, или гетерономны, и только у некоторых низших Р. они, за исключением нескольких передних и задних члеников, более или менее одинаковы и равнозначащи. У большинства Р. отдельные членики срастаются или сливаются между собой, образуя более или менее резко обособленные отделы или области тела, а именно: голову (caput), туловище (thorax) и брюшко (abdomen, фиг. 18—28). Обыкновенно у Р. голова сливается еще с туловищем, или по крайней мере с одним или несколькими члениками туловища в один отдел, так наз. головогрудь, или cephalothorax (фиг. 24—26 сth), за которой следуют оставшиеся членики туловища. У некоторых Р. слияние сегментов идет так далеко, что не только в головогруди не бывает заметно членистости, но и граница между туловищем и брюшком становится неясной, или даже членистость тела исчезает вовсе (фиг. 5, 6, 17 и 90). Вообще относительно формы тела среди Р. различных отрядов царит большое разнообразие. У многих Р. на спинной стороне вырастает еще складка кожи, которая, разрастаясь и свешиваясь вниз на брюшную сторону, прикрывает наподобие щита, или панциря (фиг. 3, 8, 9—11), или двустворчатой раковины большую или меньшую часть тела, как, например, у некоторых веслоногих (Сореpoda, фиг. 2), листоногих (Phyllopoda, фиг. 8—9), ракушковых (Ortracoda, фиг. 12) и десятиногих (Decapoda). У большинства усоногих (Cirripedia, фиг. 13—16) эта складка обрастает тело совершенно наподобие мантии, в которой отлагаются еще известковые пластинки и тем самым довершается сходство с раковиной моллюсков. С другой стороны, у некоторых паразитических Р. наблюдается вследствие образа их жизни регресс, выражающийся в полной утрате членистости, так что тело их напоминает червей, как напр. Siphonostoma (фиг. 4—6) отряда веслоногих (Copepod a) или Rhizocepha1а (фиг. 17) отряда усоногих (Cirripedia): у последних тело имеет вид мешка, снабженного на одном конце корневидными отростками, которые врастают в тело хозяина и служат паразиту для питания, происходящего эндосмотическим путем.
РАКООБРАЗНЫЕ I.

РАКООБРАЗНЫЕ II.

РАКООБРАЗНЫЕ III.
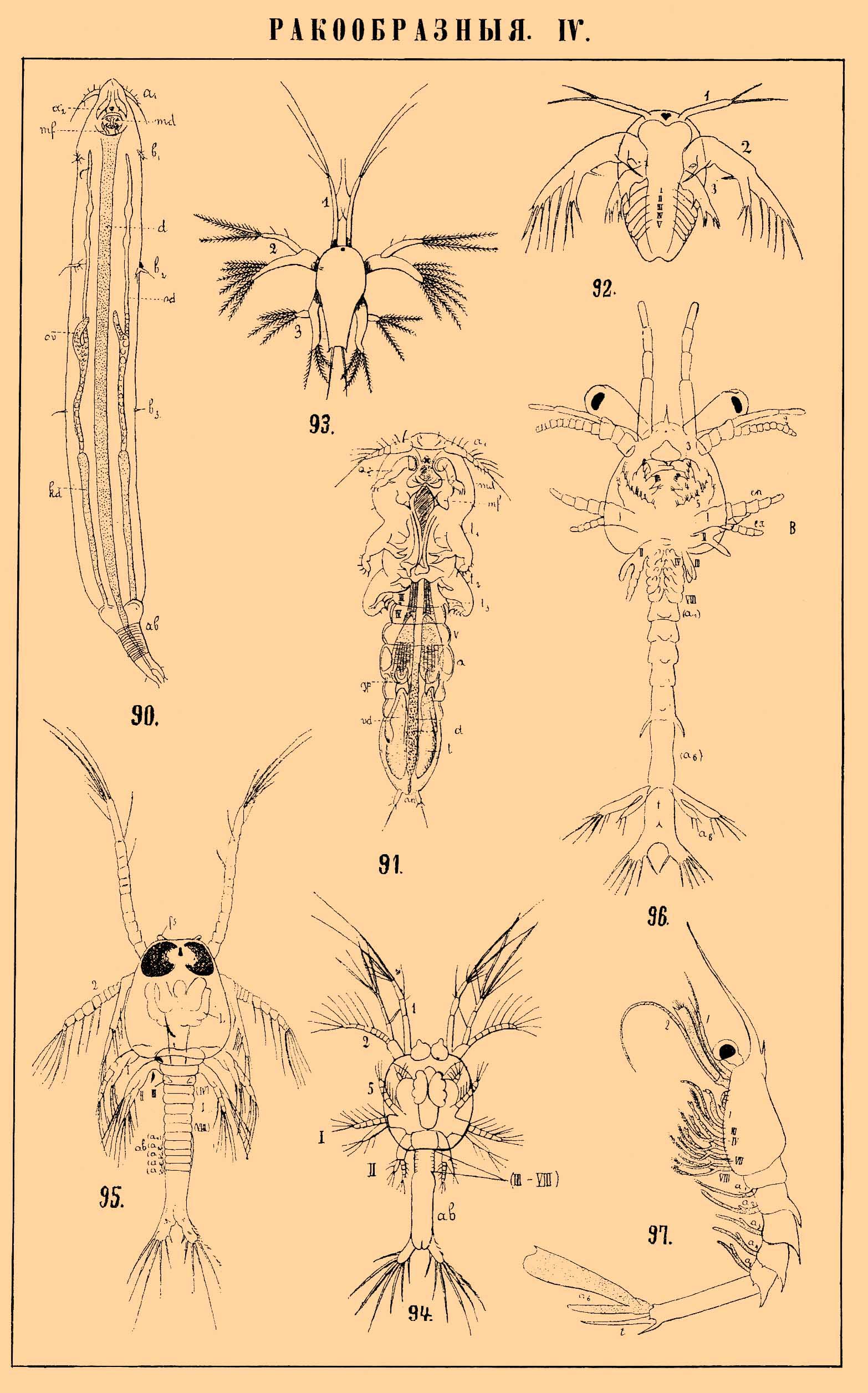
РАКООБРАЗНЫЕ IV.
К таблицам РАКООБРАЗНЫЕ
1. Cyclops canthocarpoides Fisch. (веслоногие). Самка. an1 — усики; о — глаз; so — яйца; w — яичники; а — порошица.
2. Canthocamptus staphylinus Jur. (веслоногие). Самец. аn 1 и а n2 — усики; р 1— р 4 — туловищные ножки; te — семянники; оg — половое отверстие; s — расширение vas deferens; l — верхняя губа.
3. Argulus foliaceus (веслоногие). Самка с брюшной стороны. ov — яичники; ос — глаз; р 1 — р 4 — туловищные ножки; l — хвостовой плавник; z — присоска.
4. Lamproglena pulchella Nordm. (веслоногие) с брюшной стороны; an1 — an2 — усики; рn 1 — рn 4 — туловищные ножки.
5. Chondracanthus triglae Nordm. (веслоногие). Самка.
6. Penella sagitta L. (веслоногие). Самка.
7. Branchipus stagnalis L. (жаберноногие). Самец сбоку; аn 1 и аn 2 — усики; ар — придаток; х — непарный лобный орган; h — печень; md — жвалы; gl — железка; с — сердце; in — кишечник; te — семянники; ре — penis.
8. Apus cancriformis Schä ff. (жаберноногие). Самка с брюшной стороны.
9. Apus cancriformis Schä ff. (жаберноногие). Самец со спинной стороны.
10. Daphnia similis (ветвистоусые). Самка; а 1— а 2 — усики; l — печень; au — глаз; d — кишечник; sd — раковинная железка; h — сердце; bru — выводковая камера; ab — брюшко; br — жаберные мешочки; f1—f5 — туловищные ножки; g — головной ганглий.
11. Simocephalus vetulus О. F. M ü ll. (ветвистоусые). Самка. аn 1 — an2 — усики; ос — глаз;
gа — головной ганглий; p — туловищные ноги; pа — брюшко; in — кишечник; с — сердце; va — двустворчатая раковина; х — место прикрепления к раковине.
12. Cypris fasciata (ракушковые). Самка. an1, an2 усики; ос — глаз; ab — брюшко; ov — яичник; h печень; md — жвало; m — челюсть; gl — раковинная железа.
13. Lepas anatifera (усоногие), сбоку; известковые пластинки раковины; с — саrina; t — tergum; s — scutum. — p — стебель, или ножка.
14. Lepas anatifera (усоногие). Правая половина раковины и покровы удалены; m — замыкающий мускул; l — печень; ovd — яйцевод; ov — яичник; cd — цементная железа; а' — усик; t — семянник; vd — vas deferens; p — penis.
15. Balanus Hameri As c. (усоногие).
16. Balanus tintinnabulum L. (усоногие); известковые пластинки удалены с одной стороны; s — scutum; t — tergum; o — перепонка крышки; r — rostrum; a, h, r — мышцы; l — верхняя губа; g — слепые мешки яичника; f — кожный мешек, в котором помещается тело животного; d — отверстие слухового мешка; cr — cirri, видоизмененные конечности.
17. Sacculina carcini (усоногие), паразитирующая на крабе; br — жаберная область; l — область печени и d — кишечника хозяина (краба); ks — туловище паразита; p — его ножка; mb — основная перепонка, от которой отходят корневидные отростки.
18. Nebalia Geoffroyi M. Edw. (Leptostraç a). Самка; an1 — an2 — усики; ос — глаз; ра — щупики первой пары челюстей; pe1 — pe6 — бpюшные ножки; vе — жвательный желудок; in — кишечник.
19. Gammarus Kietlinskii Dyb. (амфипода).
20. Orchestia cavimana (амфипода). Самец. с+1 — голова с 1 грудн. сегментом; II—VIII — отдельн. грудн. сегменты с конечностями; p1 — p7 — брюшные сегменты; а 1 — а 2 — усики; а — глаз; ос — пищевод; kf — челюстн. нога; br — жабры; bm — нервная цепь; g — головной ганглий; aoa — передн. аорта; sm — желудок; ud — слепая кишка; оd — яичник; h — сердце; t — семянник; vs — семянной пузырь; de — семяизвергательный канал; ehd — отверстге мочевого пузыря hd в кишечник; аор — задняя аорта; l — печеночн. придатки; ed — задняя кишка.
21. Caprella acutifrons (амфипода). Самец А сбоку; В со спинной стороны, а 1 — а 2 — усики; b2 — b8 — грудные ножки; br — жабры (вместо 4 и 5 грудн. ножки). cth+I+II — головогрудь с 2 грудн. сегментами; III—VIII свободн. грудн. сегменты; ab — рудиментарное брюшко.
22. Idothea entomon L. (равноногие) с брюшной стороны.
23. Porcellio scaber (равноногие) Latr. — 23a. То же: самка с брюшной стороны, выводковая камера наполнена яйцами.
24. Diastylis stygia (Cumacea). Самец. a1 — а 2 — усики; cth — головогрудный щит; IV — VIII — грудные сегменты; ab1 — ab7 — брюшные сегменты; p1 — p6 — брюшные ножки; en — эндоподит; еx — экзоподит.
25. Squilla (ротоногие) сбоку, cth — головогрудный щит; VI, VII, VIII — 3 задн. свободн. грудн. сегмента; a1 — а 7, — брюшные сегменты; au — глаз; а 1 — а 2 — усики; 1 — 8 — грудные ножки; 1 — 3 — челюстные ноги; p1 — p6 брюшные ножки; br — жабры на экзоподитах; р — penis.
26. Siriella thompsonii (расщепленогие). Самец. cth — головогрудный щит; VII — VIII — свободн. грудные сегменты; аb 1 — аb 7 — брюшные сегменты; p1 — p6 — брюшные ноги; br — жабры.
27. Euphausia Mülleri (расщепленогие) Claus. Самец сбоку.
28. Lophogaster typicus (расщепленогие) Sars. Самец сбоку.
29. Передние усики (antennulae) речного рака (Astacus); о — слуховой пузырь.
30. То же Cyclops serrulatus (веслоногие); rf — обонятельные волоски.
31. Вторая пара (задняя) усиков (antennae) Pseudocalanus elongatus (веслоногие).
32. То же Trebius caudatus (паразитическ. веслоногие).
33. То же Eulimnadia Agassizii (листоногие).
34. То же Euphausia pellucida (расщепленогие).
35. Жвалы (mandibulae) Notodelphys Allmannii (веслоногие); I — проксимальный, II — дистальный членник протоподита; еx — экзоподит; en — энтоподит (щупик); k — жевательная пластинка.
36. То же Cyclops tenuicornis (веслоногие).
37. То же Apus lucasanus (жаберноногие).
38. То же речного рака (Astacus); значение букв см. фиг. 35.
39. Первая пара челюстей (1 mахillае) Notodelphys agilis (веслоногие); ех — экзоподит; en — энтоподит; k — жевательная пластинка.
40. То же Euphausia pellucida (расщепленогие); значение букв то же, что в фиг. 39; ki — внутренняя жеват. пластинка, k 2 — наружная жеват. пластинка.
41. То же речного рака (Astacus).
42. Вторая пара челюстей (2 maxillae) Eucopia australis (расщепленогие); I — проксимальный, II — дистальный членик протоподита: ki — внутренняя, ke — наружная пластинка; en — энтоподит (щупик); еx — экзоподит (пластинка).
43. То же Paranebalia longipes (Leptostraca);
значение букв см. фиг. 42.
44. То же речного рака (Astacus); I — проксимальный, II — дистальный членик протоподита: ki — внутренняя, ke — наружная пластинка; en — энтоподит (щупик); еx — экзоподит (пластинка).
45. Поперечный разрез чрез тело Apus (жаберноногие) в области 7 или 8 пары ножек; h — сердце; d — кишечник; ov — яичники: bm — брюшная нервная цепочка; ah — дыхательная полость между раковиной s и телом; 1 — 6 — эндиты; br — жабры; еx — дыхательная пластинка.
46. Вторая торакальная ножка самки Daphnia similis (ветвистоусые); 1 — 5 — эндиты; еx — дыхательная пластинка, т. е. экзоподит; br — жабры.
47. То же Limnocythere incisa (ракушковые).
48. Первая торакальная ножка самца Apus longicaudatus (жаберноногие); 1 — 5 — эндиты; еx — дыхательная пластинка, т. е. экзоподит; br — жабры.
49. Плавательная ножка самки Doropygus porcicauda (веслоногие); I—II — членики протоподита.
50. Листовидная грудная ножка Nеbаliа (Leptostraca). I — II — членики протоподита; en — энтоподит; ех — экзоподит; ер — эпиподит.
51. Первая пара грудных (челюстных) ног Amphitoë penicilata (Ampbipoda). I — II — членики протоподита; 1 — 5 — членики энтоподита; k1 k1 — жевательн. пластинки; en — энтоподит.
52. Первая грудная нога Apseudes (равноногие); I — II — членики протоподита; 1 — 5 — членики энтоподита; k1 k1 — жевательн. пластинки; en — энтоподит; ер — эпиподит.
53. Вторая грудная нога Apseudes (равноногие, Amsopoda); I — II — членики протоподита; 1 — 5 — членики энтоподита; k1 k1 — жевательн. пластинки; en — энтоподит; ех — экзоподит.
54. Третья грудная нога Asellus (равноногие); I — II — членики протоподита; 1 — 5 — членики энтоподита; k1 k1 — жевательн. пластинки; en — энтоподит.
55. Первая грудная нога Asellus (равноногие); I — II — членики протоподита; 1 — 5 — членики энтоподита; k1 k1 — жевательн. пластинки; en — энтоподит; ер — эпиподит.
56. Поперечный разрез через туловище Corophium longicorne (Amphipoda); d — кишечник; h — сердце; bm — брюшная нервная цепочка; l — печень; br — жабры; brl — выводковая пластинка; ov — яйца в выводковой камере; bf — грудные ножки.
57. Первая грудная нога самки Diastylis stygia (Cumacea); ep — эниподиальная пластинка; br — жабра; en — энтоподит; еx; — экзоподит.
58. Вторая грудная нога того же.
59. Шестая грудная нога того же; brp — выводковая пластинка.
60. Вторая челюстная нога личинки Squilla (ротоногие); еn — энтоподит; ex — экзоподит.
61. Одна из трех грудных ног той же.
62. Первая грудная (челюстная) нога Lophogaster (расщепленогие); I, II — членики протоподита; 1 — 5 членики энтоподита; еn — энтоподит; ex — экзоподит; ер — эпиподит (пластинка).
63. То же Mysis (расщепленогие); I, II — членики протоподита; 1 — 5 членики энтоподита; еn — энтоподит; ex — экзоподит; ер — эпиподит (пластинка); k — жевательная пластинка.
64. Третья грудная нога Mysis.
65. Первая грудная нога Thysanoessa gregaria; I, II — членики протоподита; 1 — 5 членики энтоподита; еn — энтоподит; ex — экзоподит; ер — эпиподит (пластинка); br — жабры.
66. Конец брюшка Diastylis stygia (Cumaсеа); V, VI, VII — членики брюшка; VII — telson; p6 — шестая пара брюшных ног; en — энтоподит; ex — экзоподит.
67. Вторая брюшная нога Siriella Thompsonii (расщепленогие) с жаброй, экзо- и энтоподитом.
67а. Конец брюшка Siriella; VI — VII — абдоминальн. сегменты; еn — эндоподит; ех — экзоподит; g — слуховой орган.
68. Хвостовой плавник Anilocra (равноногие); VI — шестой брюшной членик; VII — telson; p6 — шестая пара брюшных ног с экзоподитом ех и энтоподитом еn.
69. Конец брюшка Lysianassa producta (Amphipoda) с 4, 5 и 6 брюшной ногой р 4, p5, p6, IV—VII — брюшные сегменты; en — энтоподит; ех — экзоподит.
70. Схема, поясняющая механизм движения членистого тела ракообразных, тело в вытянутом состоянии; ct — больший сегмент, за ним 4 меньших; наружный скелет обозначен темными линиями, а сочленовые кожицы пунктиром; в а сегменты соприкасаются между собой (неподвижные точки рычагов); t — спинной скелет, s — брюшной, d — спинные мышцы; v — брюшные; tg -спинные сочленовые кожицы; vg — брюшные.
71. Схема, поясняющая механизм движения членистого тела ракообразных, тело в вытянутом состоянии; ct — больший сегмент, за ним 4 меньших; наружный скелет обозначен темными линиями, а сочленовые кожицы пунктиром; в а сегменты соприкасаются между собой (неподвижные точки рычагов); t — спинной скелет, s — брюшной, d — спинные мышцы; v — брюшные; tg -спинные сочленовые кожицы; vg — брюшные. Тело согнуто на брюшную сторону вследствие сокращения брюшных мышц v.
72. Кишечник Asellus aquaticus (равноногие); oe — пищевод, km — жевательный желудок, d — средняя кишка, a — задняя кишка, l — печеночные придатки.
73. Нервная система Limnadia (листоногие), передняя часть; ua — нерв непарного глаза; au — нервы парных глаз; а 1 — а 2 — нервы первой и второй пары усиков; g — головной мозг; а 2g — ганглий второго усика; sc — окологлоточная комиссура; у — подглоточная комиссура.
74. Нервная система Apseudes Latreillii (равноногие, Anisopoda); md — мандибулярный ганглий; mх 1 — mх 2 — ганглий 1-й и 2-й пары челюстей; I—VIII — торакальные ганглии; 1—6 — абдоминальные ганглии; ua — нерв непарного глаза; au — нервы парных глаз; а 1 — а 2 — нервы первой и второй пары усиков; g — головной мозг; а 2g — ганглий второго усика; sc — окологлоточная комиссура; у — подглоточная комиссура.
75. Нервная система Euphausia pellucida (расщепленогие); md — мандибулярный ганглий; mх 1 — mх 2 — ганглий 1-й и 2-й пары челюстей; I—VIII — торакальные ганглии; 1—6 — абдоминальные ганглии; ua — нерв непарного глаза; au — нервы парных глаз; а 1 — а 2 — нервы первой и второй пары усиков; g — головной мозг; а 2g — ганглий второго усика; sc — окологлоточная комиссура; у — подглоточная комиссура.
76. Нервная система Astacus fluviatilis (десятиногие); s — симпатическая нервная система; сg — ганглии комиссур; bg — подглоточный ганглий; значение остальных букв см. фиг. 73 и 74.
77. Нервная система Maja squinado (десятиногие); m — желудок; sg — ганглии симпатической нервной системы; значение остальных букв см. фиг. 73 и 74.
78. 2 обонятельные трубки Nebalia (Leptostraca).
79. Перистая чувствительная щетинка (слуховой волосок) Apseudes (равноногие).
80. Осязательный волосок (tb) Branchipus (листоногие — жаберноногие); с — кутикула; hy — гиподермическая клетка; gz — ганглиозная клетка; n — нервное волокно.
81. Представитель равноногих сбоку; аn 1 — передние, аn 2 — задние усики; ctb — головогрудь; II—VIII — 7 свободных грудных сегментов; a1 — a7 — 7 брюшных сегментов; br — жабры; g — головной мозг; d — кишечник; h — сердце; о — отверстие (ostium) сердца; рс — перикардий; va — передняя аорта; la — боковые артерии; t — грудные артерии; ha — печеночные артерии; si — боковые синусы грудной области; sa — брюшной синус; abg — место прикрепления грудных ног; рg — субнейральный сосуд; bp — жаберно-перикардиальный сосуд; zg — приводящие жаберные сосуды; аа — брюшная аорта; os — отверстия боковых синусов.
82. Поперечный разрез брюшка Conilera cylindracea (равноногие); b — сердце; x, у — мускульные пластинки, служащие для двигания жабер (плеоподы); ara — брюшные артерии мускульных пластинок; еx — внешняя, en — внутренняя жаберная пластинка (экзо- и энтоподиты); аg — отводящий, zg — приводящий жаберный сосуд; bm — нервная цепочка; рn — субнейральный сосуд; sa — брюшной синус.
83. Представитель амфипод сбоку; pt — перикардиальные сосуды, отходящие от эпимер ер, конечностей brf и жабер в груди; ра — перикардиальные сосуды брюшка; s — брюшной синус; ер — эпимеры; рl — плеоподы; остальные буквы см. фиг. 81.
84. Поперечный разрез через головогрудь речного рака (Astacus) в области сердца; kd — жаберная крышка; k — жабры; kh — дыхательная, или жаберная, полость; ер — боковая стенка головогруди; рс — перикардий; h — сердце; sa — arteria sternalis; l — печень (hераtoраnсrеas); d — кишечник; abm — брюшные, dbm — спинные мышцы брюшка; bm — нервная цепь; sn — субкейральный сосуд; bf — ходильная нога; vs — брюшной синус; ov — оварий; стрелы указывают направление движения крови.
85. Поперечный разрез чрез головогрудь Birgus latro (Decapoda Anomura) в области сердца; kd — жаберная или легочная крышка; h — сердце; k — жабры; ah — дыхательная полость; p — перекардий; ek — жаберн. кровеносн. сосуды, идущие к сердцу; а 1 а 2 а 3 а 4 — кровеносные сосуды, идущие от сердца; lb — легочный пучок; еl — кровеносный сосуд, идущий от легких к сердцу; еl 1 — вблизи окончания в перикардий.
86. Половые органы самки Lernanthropus (веслоногие); ov — яичники; od — яйцеводы; kt — железки; rs — семяприемник; go — половое отверстие; p — канал, ведущий к сперматофору; bk — бурое тело; sp — сперматофора.
87. Половые органы самца Lernanthropus (веслоногие); t — семенник; vd — семяпровод; st — сперматофорный мешок; go — половое отверстие.
88. Половые органы самки Asellus aquaticus (равноногие); ov — яичники; od — яйцеводы.
89. Половые органы самца Asellus aquaticus (равноногие); t — семенники; vd — семяпроводы; pl — 1 и 2 пара брюшных ног.
90. Самка Lernaeascus nematoxys (веслоногие); а 1 — передн1е, а 2 — задние усики; md — жвалы; mf — челюстные ножки; b 1 b2 b3 — грудные ножки; ov — яичник; od — яйцевод; kd — железка; аb — брюшко; d — кишечник.
91. Самец Lernaeascus nematoxys (веслоногие); t1 t2 t3 — грудные ножки; III, IV, V — грудные сегменты; a1 — 1 брюшной сегмент; gр — половая пластинка; t — семенник; vd — семяпровод; остальные буквы см. фиг. 90.
92. Личинка науплиус Apus'a (листоногие) с зачатком 5 передних туловищных сегментов.
93. Личинка науплиус Penaeus'a (десятиногие); 1—3 — конечности головы.
94. Личинка протозоэа Penaeusa; III—VIII — зачатки 3—8 туловищных сегментов; ab — брюшко; 1—5 — конечности головы; I, II — конечности груди.
95. Личинка протозоэа Penaeusa, более взрослая стадия со спинной стороны; fs — фронтальный (лобный) орган чувств; L — печень; ab — брюшко; IV—VIII — грудные сегменты, а 1 — а 2 — абдоминальные сегменты.
96. Личинка зоэа Penaeus'a; en — энтоподит; ех — экзоподит; t — тельзон; остальные буквы см. фиг. 95.
97. Личинка, так назыв. мизидная стадия, Penaeus'a. сбоку.
Конечности. Каждый членик, входящий в состав тела Р., за исключением последнего, бывает снабжен на брюшной стороне одной парой членистых отростков или конечностей, соответствующих параподиям кольчатых червей (см.). Представляя более высокую степень дифференцировки, конечности Р. отличаются от последних тем, что 1) сочленены с телом помощью особых суставов, 2) состоят из отдельных, сочлененных между собой (членистых) частей и 3) имеют самостоятельную мускулатуру, являясь таким образом подвижными рычагами. Эти конечности выполняют различные функции и служат для плавания, хождения, прикрепления, захвата и измельчения (жевания) пищи, осязания, совокупления и носки яиц. Исполняя ту или другую из перечиссленных функций, конечности изменяются морфологически и весьма разнообразны по своему строению не только у различных представителей Р., но и в разных члениках одного неделимого. Первые две пары конечностей, усики, или antennae, помещаются у Р. в предротовом отделе тела и представляют из себя конечности первых двух сегментов, переместившихся во время эмбрионального развития вперед и иннервируемых от глоточной комиссуры. Таким образом, усики Р. существенно отличаются от усиков насекомых (см.), соответствующих первичным щупальцам кольчатых червей (см.). Первая пара усиков Р. может быть противопоставлена всем прочим конечностям, так как состоит лишь из одного ряда члеников, хотя и имеет иногда 1—3 вторичных придатков или жгутов (фиг. 1, 2, 4, 7, 10—12, 14, 18—21, 2 4, 25, 29, 30). Они служат по преимуществу органами осязания, но являются носителями и органов обоняния (фиг. 30), а подчас и слуха (фиг. 29). У некоторых ракушковых (фиг. 12) и веслоногих (ф. 10) они служат еще и для плавания. Все остальные конечности состоят из двух ветвей или двух рядов члеников и представляют так наз. расщепленные ножки, составляющие характерную особенность класса Р. К этим расщепленным ножкам, считаемым за примитивную и родоначальную форму конечностей, могут быть сведены все конечности всех Р., как бы разнообразно они ни были устроены. В расщепленной ножке различают три части: 1) основную часть, или ствол ноги (protopodit); 2) внутреннюю ветвь (endopodit) и 3) наружную ветвь (exopodit). Ствол ноги состоит из двух члеников, из коих проксимальный сочленен с сегментом тела, а дистальный несет обе ветви; каждая ветвь состоит из ряда члеников, причем внутренняя обращена к медианной плоскости тела, а наружная — в противоположную сторону. В разных конечностях, соответственно той или другой функции их и у различных Р., эти три части, или отдела, представляют неодинаковое развитие, а частью и совершенно исчезают, обусловливая различную форму конечностей. Вторая пара усиков, или вторая пара (фиг. 31—34) конечностей, состоит у большинства Р. из двух рядов члеников и является по преимуществу органами осязания; лишь у низших Р. они служат для плавания (у водяных блох среди листоногих и у ракушковых), для захвата добычи (у жаброногих) и для прицепления (у паразитических веслоногих); у некоторых Р. они рудиментарны (некотор. амфипода и равноногие) или отсутствуют вовсе (у усоногих). Третья пара конечностей (фиг. 35—38), или жвалы (mandibulae), расположена впереди и с боков ротового отверстия и служит для измельчения пищи. Они первоначально являются типичными расщепленными ножками (и остаются таковыми у веслоногих и ракушковых), которые, принимая на себя жевательную функцию, изменяются многосторонне. Проксимальный, или основной, членик ствола (протоподита) превращается в твердую пластинку, зазубренную на своем свободном, обращенном ко рту краю, тогда как остальные части конечности остаются рудиментарными в виде маленького щупика (жвальное щупальце) или исчезают вовсе. Четвертая пара конечностей (фиг. 39—41), или передняя пара челюстей (1 maxillae), расположена вблизи рта и служит, подобно 3-й и 5-й паре конечностей, для жевания. Они имеют вид нежных расщепленных пластинок, в которых можно различить основной ствол (protopodit), снабженный жевательными лопастями, а также и части, соответствующие обеим ветвям; таким образом, в них еще лучше, чем в жвалах, сохраняется тип расщепленных ножек. Пятая пара конечностей (фиг. 42—44), или задняя пара челюстей (2 maxillae), построена по типу предыдущей, но еще более напоминает расщепленные ножки, так как сохраняется ее наружная ветвь (exopodit); лишь у усоногих задняя пара челюстей мала и рудиментарна. Эти пять пар конечностей, прикрепляющиеся к первым пяти сегментам, называются головными конечностями, так как голова Р. образуется слиянием первых пяти сегментов. Следующие за ними конечности называются конечностями туловища; число их у низших Р., или Энтомострак, изменчиво и постоянно у высших Р. или Малакострак. У последних (фиг. 18—28, 81 и 83) согласно подразделению туловища на грудь и брюшко различают 8 пар грудных конечностей и 6 пар брюшных, или абдоминальных, конечностей. Туловищные конечности Энтомострак (фиг. 2—4, 7—8, 10—12, 14, 16, 46—49) соответствуют грудным конечностям Малакострак и бывают чрезвычайно разнообразно устроены. Так, у жаброногих отряда листоногих встречаются 10—36 пар конечностей, имеющих вид листообразных или пластинчатых плавательных ножек с жаберными придатками и служащих помимо того еще для привлечения пищи. Каждая ножка (фиг. 46—49) состоит из основного ствола (protopodit), несущего на внутренней стороне 6 придатков или лопастей (эндитов), на внешней стороне сплющенную дыхательную пластинку (exopodit) и мешкообразный жаберный придаток (epipodit). У ветвистоусых отряда листоногих всего 4—6 пар туловищных конечностей, а у ракушковых даже 2 пары, имеющие вид длинных многочленистых ног без дыхательной пластинки и жаберного придатка. Туловищные конечности (фиг. 2) веслоногих (4—5 пар) служат для плавания и являются типичными расщепленными ножками, тогда как у усоногих (3—6 пар) они служат для привлечения добычи (ф. 14) и состоят из основного отдела (protopodit), двух длинных и тонких многочленистых жгутообразных ветвей (exopodit и endopodit), густо усаженных волосками, и называются усоножками. Туловищные конечности Leptostr a ca (фиг. 18 и 50) представляют переход от конечностей низших Р. — Entomostraca к высшим P. — Malacostraca. У них различают 8 пар грудных и 6 пар брюшных конечностей; первые все одинаковы и являются пластинчатыми образованиями, напоминающими конечности листоногих, и состоят (фиг. 50) из двучленистого основного отдела (protopodit), несущего пластинчатый жаберный придаток (epipodit), жаберную пластинку (exopodit) и пятичленистый щупик (endopodit); из 6 пар брюшных конечностей первые 4 пары служат для плавания и являются типичными расщепленными ножками, напоминающими конечности веслоногих, тогда как 2 задние пары представляются рудиментарными, в виде коротких одно- или двучленистых придатков. У высших Р. — Malacostraca (фиг. 18—28, 50—65, 81 и 83) встречающиеся всегда в количестве 8 пар грудные конечности весьма разнообразны как по функции, так и по наружному виду. Подобно тому, как в разных отрядах Малакострак различное число грудных сегментов может соединяться или сливаться с головным отделом, точно так же и различное количество передних грудных конечностей (1—5 пар) придвигается ко рту; они являются вспомогательными органами для принятия пищи. В силу этой функции они изменяются морфологически и в своем строении представляют переход от челюстей (max i llae) к собственно ножкам (pedes) и называются челюстными ножками (pedes maxillares). Исходной формой для грудных конечностей Малакострак является описанная конечность Leptostraca (фиг. 50); к основному стволу (protopodit) конечности Малакострак прикрепляются epipodit, exopodit и пятичленистый endopodit. Очень часто последние два придатка атрофируются, и тогда конечность является в виде нерасщепленной ножки, состоящей из расположенных в один ряд 7 члеников. У членистогрудых раков (Arthrostraca) имеется только одна пара челюстных и 7 пар плавательных или ходильных ног (фиг. 51—65); причем на средних ножках у самок имеются особые пластинки, которые, налегая черепицеобразно друг на друга, замыкают под брюшной стенкой тела пространство (зародышевая камера), куда откладываются яйца (фиг. 56) и где развиваются из них зародыши. У амфипод (фиг. 56) к основному членику средних и задних ножек с внутренней стороны прикрепляются нежные жаберные мешочки (epipodit) по одному к каждой ножке, тогда как у другого отряда членистогрудых раков — равноногих (Isopoda, фиг. 52—65) жаберные пластинки помещаются лишь на челюстных ножках, а вторая пара грудных ножек (ф. 53) у некоторых представителей снабжена клешней. Не менее разнообразны грудные конечности щитогрудых раков (Th o racostraca). Так, у отряда Cumacea (фиг. 24, 57—59) имеется одна пара челюстных ножек, снабженных жаберными придатками, а к 2 и 6 паре грудных ножек у самок прикреплены пластинки, прикрывающие зародышевую камеру. У отряда ротоногих (Stomatopoda, фиг. 25, 60—61) 5 пар передних конечностей являются челюстными ножками, которые, кроме того, служат еще для захвата добычи и несут пластинчатые жаберные придатки; 3 пары задних грудных конечностей — типичные расщепленные ножки и служат для хождения (ходильные ножки). Грудные конечности расщепленогих (Schizopoda, фиг. 26—28, 62—65) по своему строению, с одной стороны, примыкают к грудным конечностям Leptostraca, с другой же стороны, представляют переход к грудным конечностям высших Р., а именно к десятиногим ракам (Decapoda). Все 8 пар грудных конечностей довольно однородны и представляют типичные расщепленные ножки: у сем. Euphausidae (фиг. 27) на 2 передних парах конечностей имеются зачатки жевательных пластинок, а у семейства Mysidae (фиг. 63—64) — две пары типичных челюстных ножек. Наконец, у десятиногих раков (Decapoda) имеются 3 пары челюстных ножек, служащих только для принятия пищи, и 5 пар ходильных ног, из коих передние пары часто снабжены клешнями, особенно сильно развитыми на первой паре ходильных ног (относительно строения конечностей Decapoda (см. Десятиногие раки и таблицу рисунков). Брюшные конечности, или абдоминальные ножки, или плеоподы малакострак (фиг. 18—28, 66—69, 81 и 83) встречаются постоянно в количестве 6 пар, причем последний (седьмой) сегмент тела, называемый также тельзоном, всегда лишен конечностей. У членистогрудых раков (Arthrostraca) они представляются типичными расщепленными ножками; у амфипод (фиг. 19—21, 69 и 83) 3 передние пары обращены вперед, хорошо развиты и служат для плавания, а три задние пары обращены назад, короче и служат часто для прыгания; у равноногих (Isopoda, фиг. 22—23, 68 и 81) они пластинчаты и служат для дыхания, а последняя пара (у плавающих, морских форм) вместе с последним сегментом образует большой плавник или же (у наземных форм, напр. мокрица) имеет форму стилета. У щитогрудых раков (Thoracostraca) абдоминальные конечности являются также расщепленными ножками. У ротоногих (Stomatopoda, фиг. 25) 5 передних пар служат для плавания и снабжены разветвленными жаберными придатками, а 6 пара срослась с тельзоном в сильный хвостовой плавник. У Cumacea (фиг. 24 и 66) 5 передних пар служат для плавания у самцов и отсутствуют у самок, а 6 пара вилкообразна. У расщепленогих (Schizopoda, фиг. 26—28 и 6 7) 6 пара вместе с последним сегментом образует плавник, а 5 передних пар у самцов являются плавательными ножками, причем у некоторых (Euphausidae) передние две пары приспособлены к совокуплению. У десятиногих раков (Decapoda) 6 пара плеопод также срастается с последним сегментом и образует сильный хвостовой плавник, а из 5 передних пар у самцов две первые пары являются органами совокупления, а у самок первая пара рудиментарна, тогда как 4 остальные служат для носки яиц. У короткохвостых Decapoda все плеоподы недоразвиты (см. Десятиногие раки, фиг. 1, 2, 3, 7, 8, и 12) или отчасти отсутствуют вовсе.
Покровы и мускулатура. Все тело Р. покрыто кожистым, твердым панцирем, представляющим наружный скелет и служащим, помимо защиты внутренних органов, еще для прикрепления мускулов. Панцирь покрывает со всех сторон сегменты, а между отдельными сегментами, т. е. на границе их, находится тонкая, нежная сочленовая кожица, делающая возможным подвижность тела. При сокращении и поворотах тела сегменты входят отчасти один в другой наподобие частей подзорной трубы (фиг. 70—71). Панцирь Р. состоит из хитина, т. е. рогового, органического азотсодержащего вещества, не растворимого в щелочах и кислотах, органических и минеральных; при кипячении с концентрированной серной кислотой хитин распадается на сахар и аммиак и отличается этим от других подобных образований, встречаемых в животном царстве, как напр. роговое вещество губок, конхиолин раков и моллюсков и т. под. Хитиновый панцирь Р. представляет продукт выделения эктодермального эпидермиса, или гиподермиса, или так назыв. хитиновой оболочки. Последняя очень тонка, тогда как хитиновый панцирь достигает значительной толщины и имеет слоистое строение, указывающее на постепенный прирост его. На наружных покровах в разных местах тела встречаются различные отростки, шипы, волоски и т. под., представляющие выросты хитинового слоя. В хитиновом покрове Р. откладываются еще углекислые соли извести, которые, пропитывая его, делают плотным и твердым панцирем. Благодаря этим отложениям панцирь теряет свою эластичность, делается хрупким и легко раскалывается; само присутствие панциря послужило поводом к латинскому названию Р. — Crustacea, или черепокожные. Этот панцирь сбрасывается у некоторых Р. периодически и заменяется новым, представляя процесс, известный под названием линьки. В панцире имеется много пор, а под гиподермическим слоем залегают различные железы, число, расположение и строение которых весьма разнообразно; некоторые из них заведуют выделением. Некоторые брюшные и ножные железы соответствуют коксальным железам насекомых, а у наземных равноногих (мокрица) встречаются еще паутинные железы. Мускулатура хорошо развита и состоит из отдельных мышц или мускульных тяжей с сухожилиями, которые прикрепляются к внутренней стенке наружного скелета или его отросткам или перегородкам (фиг. 70—71). В общем мускулатура очень сложна и чрезвычайно разнообразна у различных представителей Р. и, понятно, находится в тесном соотношении с общей формой тела и членистостью. Различают: 1) мышечные соединения между сегментами тела, 2) мускулатуру конечностей, 3) мышцы, соединяющие тело с конечностями. Мышцы Р. являются полосатыми и по своему гистологическому строению напоминают поперечно-полосатые мышцы позвоночных животных.
Органы пищеварения. Кишечник всех Р. состоит из трех отделов передней, средней и задней кишки, различной величины у разных отрядов и снабженных разнообразными придатками (фиг. 1—2, 4, 7, 10—12, 18, 20, 27, 72, 81, 83, 90). Ротовое отверстие помещается на брюшной стороне и бывает окружено передней и задней губой, представляющими складки кожи, и кроме того челюстями, а у высших Р. еще челюстными ножками, служащими для измельчения пищи. Рот ведет в переднюю кишку эктодермального происхождения, выстланную хитиновым слоем, который сбрасывается при линьке и заменяется новым; у низших Р. (Entomostraca) различают только один пищевод, тогда как у высших Р., Malacostraca, два отдела: восходящий в спинно-брюшном направлении пищевод и большой жевательный желудок, имеющий форму широкого мешка и в котором различают передний, больший или кардиальный отдел, снабженный хитиновыми жевательными пластинками, служащими для измельчения пищи, и задний, меньший пилорический отдел (см. Десятиногие раки, фиг. 3). Особых железок, открывающихся в переднюю кишку — нет. У некоторых десятиногих раков по бокам желудка помещаются так назыв. ротовые жерновки (см.). Передняя кишка переходит в прямую среднюю кишку эктодермального происхождения, которая у Энтомострак очень длинна, занимает большую часть тела и состоит из желудка и длинной тонкой кишки. В первый открываются печеночные придатки, встречающиеся в различном числе; они бывают разнообразно устроены, а у веслоногих (Copepoda) отсутствуют вовсе. У Малакострак средняя кишка значительно короче (в особенности у равноногих и десятиногих) и принимает одну пару двулопастных желез или печень, которую правильнее было бы назвать сообразно ее физиологической функции — hepatopancreas (фиг. 72 l). В этой железе различают ферментные клетки, выделения которых переваривают фибрин, и печеночные клетки, выделением которых крахмал переводится в сахар. Средняя кишка переходит затем в заднюю кишку эктодермального происхождения, выстланную так же, как и передняя кишка, тонким слоем хитина. Задняя кишка у Энтомострак коротка и открывается наружу через порошицу обыкновенно на спинной стороне тела; у Малакострак (особенно у десятиногих) задняя кишка очень длинна и открывается порошицей на брюшной стороне в последнем сегменте наружу. Вследствие паразитического образа жизни происходит атрофия кишечника, доходящая иногда до полного исчезновения его; у усоногих (Cirripedia) встречаются разные степени атрофии кишечника, а у корнеголовых (Rhizocephala) кишечник отсутствует даже у личинок.
Нервная система Р. построена по типу нервной системы кольчатых червей (см.) и может быть производима от нее. На основании сравнительно-анатомических и эмбриологических данных нервная система Р. сводится к следующей схеме: на спинной стороне, а именно над глоткой, в переднем конце тела помещается 1 пара надглоточных ганглиев, или головной мозг, соединенных между собой перемычкой, или комиссурой; от них отходят нервы, направляющиеся к передним усикам (antennae), глазам и другим органам чувств. Все остальные сегменты имеют по одной паре ганглиев, расположенных на брюшной стороне тела, а именно под кишечником, и соединенных между собою поперечными комиссурами, а по длине — продольными комиссурами, так что в общей сложности образуется брюшная нервная лестница, или брюшная нервная цепочка, которая соединяется с надглоточными ганглиями при помощи окологлоточной комиссуры. В каждом сегменте от каждой пары ганглиев отходят нервы, направляющиеся к внутренним органам и конечностям. В таком виде нервная система встречается лишь в эмбриональном состоянии или у немногих взрослых форм Р., тогда как у всех прочих Р. наблюдаются отклонения от описанной схемы. Все изменения и отклонения от данной схемы, встречающиеся у разных представителей различных отрядов Р., могут быть сведены к следующим факторам: 1) постепенное сближение двух ганглиев одной пары вследствие укорачивания поперечных комиссур, ведущее наконец к полному слиянию двух ганглиев в один; 2) сближение последующих ганглиев вследствие укорачивания продольных комиссур; 3) слияние и срастание некоторых последующих ганглиев и перемещение их в другие сегменты и 4) полное слияние всех брюшных ганглиев в одну брюшную массу. У низших Р. (Entomostraca), в особенности у листоногих (Phyllopoda), встречается примитивное строение нервной системы, т. е. нервная лестница (фиг. 73), причем каждая пара ганглиев соединена двумя поперечными комиссурами, тогда как в других отрядах этого подкласса имеем большую или меньшую концентрацию нервной системы. Что касается высших Р. (Malakostraса), то у Leptostraca имеется еще примитивное отношение, так как число ганглиев соответствует числу сегментов; у равноногих (Isopoda) мы имем также нервную лестницу (фиг. 74), но наблюдается концентрация и слияние ганглиев абдоминальных сегментов; у расщепленогих (Schizopoda) — мандибулярные, максиллярные и торакальные ганглии (фиг. 75 md, mх, I—VI) являются самостоятельными, хотя сближены между собой и состоят каждый из двух ганглиев и двух продольных комиссур, тогда как абдоминальные ганглии и их продольные комиссуры слились в одну брюшную нервную цепь (фиг. 75, 1—6), но места слияния их остаются заметными. У десятиногих (Decapoda) мы имеем нервную цепь, о которой по наружному виду нельзя сказать, что она произошла слиянием двух стволов (фиг. 76) и только между 6-м и 7-м торакальным ганглием имеются две комиссуры, т. е. два продольных ствола, образующих отверстие для прохождения брюшной артерии (art. sternalis). Наконец, у короткохвостых десятиногих (Brachyura) все ганглии брюшной цепи сливаются в одну нервную массу (фиг. 77). Кроме центральной нервной системы, у Малакострак встречается еще симпатическая нервная система (фиг. 76 s, 77 sg), иннервирующая органы пищеварения и кровообращения и соединяющаяся с головным мозгом при помощи непарного нерва.
Органы чувств Р., за исключением глаз, еще мало исследованы. Органы осязания (фиг. 80) помещаются на усиках, ножках и других местах тела и имеют вид маленьких волосков или щетинок, отличающихся от прочих щетинок тем, что у их основания находится обыкновенно ганглиевая клетка, которая находится в связи с прочей нервной системой.
Органы обоняния (фиг. 30 и 78) имеют вид прозрачных, нежных колбочек, нитей или мешков с тонкой хитиновой оболочкой и встречаются обыкновенно на усиках.
Органы слуха (фиг. 79) встречаются большей частью на переднем конце тела у основания усиков: они имеют форму ямочек (фиг. 16), покрытых тонкими волосками, несущими отолиты, или замкнутых пузырьков (фиг. 29 и 67 а), заключающих внутри отолиты. Кроме того, у некоторых Р. встречается еще особый парный лобный или фронтальный орган чувств (фиг. 95), помещающийся на переднем конце тела, физиологическое значение которого еще не выяснено.
Глаза Р. бывают двоякие: 1) непарный глаз, встречающийся у личинок низших Р. (Entomostraca, фиг. 93), а также и у многих взрослых Энтомострак (фиг. 1—2, 7, 10—12) и называется науплиусовым глазом; у высших Р. (Malacostraca) он встречается также в личиночном состоянии, но исчезает при дальнейшем развитии. Такой непарный глаз, как мы его встречаем у свободноживущих веслоногих (Copepoda), произошел слиянием трех глаз и состоит из пигмента и ретинальных клеток, к которым подходят нервные волокна, отходящие от оптического ганглия. 2) Парные глаза (см. Глаз, фиг. 11) встречаются главным образом у Малакострак (фиг. 18, 20, 25—28, 95—97), но попадаются и у Энтомострак (фиг. 3 и 7), иногда даже совместно с простым (непарным) глазом. Нерв, иннервирующий такой глаз, подходя к нему, образует вздутие — ganglion opticum, от которого идут нервные ветви, соединяющиеся в ganglion retinale, имеюший вид полушария, от которого радиально отходят нервы, направляющиеся к сетчатке, или ретине; последняя отделена от нервных стволиков так называемой базальной перепонкой, а сам глаз выступает наружу наподобие полушария или колбочки. Retina состоит из отдельных составных элементов, или r etinulae, из коих каждая произошла слиянием 4—7 клеток, которые в центре, т. е. месте соединения, выделяют стекловидную, сильно преломляющую свет палочку — рабдом (см.). На поверхности ретинул собран пигмент, который продолжается еще в пигментный столбик — совокупность их образует так называемую Chorioidea. Далее идет стекловидное тело, которое выделяет сильно преломляющий свет кристаллический конус, имеющий вид четырехгранной усеченной пирамиды; за этим слоем идет ряд кристаллических клеток и, наконец, прозрачная роговая оболочка, или Cornea, которая у высших Р. соответственно каждой отдельной ретинуле имеет форму двояковыпуклой линзы, или фасетки — совокупность их образует сложный, или фасетированный, глаз. В прежнее время полагали, что изображение получается в каждом глазе особое или, иными словами, получается множество изображений. В настоящее время эту теорию оставили, заменив ее так называемой теорией мозаичного видения, в силу которой такой сложный глаз должен давать прямое изображение предмета, отдельные части или точки которого образуются отдельными глазами.
Кровеносная система. У Р., как вообще у всех суставчатоногих, нет замкнутой кровеносной системы; снабженные собственными стенками кровеносные сосуды находятся в непосредственном сообщении с кровеносными лакунами или синусами. Последние не имеют особых (собственных) стенок, а образуют лишь щели или полости между отдельными органами и представляют, таким образом, полость тела. На основании современных данных сравнительной анатомии и эмбриологии весьма вероятно, что кровеносная система родоначальной формы Р. была устроена следуюшим образом: на спинной стороне тела, над кишечником, помещался длинный, трубкообразный, сократимый сосуд, или сердце, занимавший всю длину тела. В каждом сегменте этот сосуд снабжен одной парой отверстий, или остий, помещающихся сбоку и через которые содержимое трубки, т. е. кровь, находится в сообщении с окружающим спинной сосуд синусом, представляющим часть полости тела (перикардиальный синус). Кровь поступает из перикардиального синуса чрез заднее отверстие и парные остии в спинной сосуд, передвигается в нем сзади наперед и выходит через переднее отверстие, попадая в лакунарную систему, находящуюся под кишечником и идущую в конечности; окисляясь в жабрах, кровь движется в брюшных лакунах в обратном направлении, т. е. спереди назад, и снова возвращается в перикардиальный синус. По такой схеме устроена кровеносная система низших Р., причем она ближе всего подходит к данной схеме у представителей листоногих жаберноногих (P hyllopoda Branchiopoda). У прочих Энтомострак сердце имеет форму мешка или пузыря (фиг. 10), снабжено лишь одной парой остий и переходит на переднем конце в короткий сосуд. Такая кровеносная система встречается у ветвистоусых (Сlаdocera), некоторых веслоногих (Copepoda) и немногих ракушковых (Ostracoda). У всех прочих Энтомострак кровеносной системы нет, т. е. вся полость тела наполнена кровью, причем это явление не примитивное, а вторичное. У промежуточных Р., т. е. у Leptostraca, кровеносная система напоминает кровеносную систему листоногих, но в то же время представляет и переход к кровеносной системе высших Р. Сердце является трубчатым, но число остий, снабженных еще к тому клапанами, ограничено; сердце на переднем и заднем конце продолжается в переднюю и заднюю аорту (с клапанами), открывающиеся в брюшной синус, отделенный перегородкой от спинного, или перикардиального, синуса. У высших Р. (Malacostraca), а именно у членистогрудых (Arthrostraса), сердце трубчатое и помещается у Аmphipoda в торакальном отделе (фиг. 20 и 83), а у равноногих (Isopoda) в абдоминальном (фиг. 81); от сердца отходит аорта и артерии, переходящие в лакунарную систему, идущую в жабры и сообщающуюся с перикардиальным синусом. Из щитогрудых (Thoracostraca) y ротоногих (Stomatopoda) сердце трубчатое, тогда как у других отрядов укорочено и мешкообразно; другой характерный признак этой группы Р. — присутствие так называемой брюшной или subneural'ной артериальной системы (фиг. 84), помещающейся на брюшной стороне под нервной цепочкой и сообщающейся с сердцем при помощи брюшной артерии (art. sternalis), идущей в спинно-брюшном направлении и проходящей между 6 и 7 (фиг. 76) торакальным ганглием (относительно органов кровообращения высших Р., т. е. десятиногих, см. Десятиногие, табл. фиг. 3). Кровь Р. представляется обыкновенно бесцветной или желтоватой, зеленоватой или красноватой жидкостью, в которой плавают амебоидные кровяные тельца. Красноватый цвет крови обусловливается присутствием гемоглобина.
Органы дыхания находятся в зависимости и в тесной связи с органами кровообращения. У громадного большинства Р. встречаются жабры, и только у некоторых равноногих (Isopoda), ведущих наземный образ жизни, встречаются воздухоносные трубочки, которые являются измененными жабрами, а по функции соответствуют трахеям насекомых (см.). Устройство и расположение жабр представляют важный систематический признак для Р. Они располагаются всегда на таких местах тела, которые являются особенно благоприятными для постоянного обмена воды, т. е. на конечностях, и у листоногих (Phyllopoda) имеют вид маленьких мешочков или пластинок (фиг. 7 и 8) помещающихся между двумя ветвями (ехоpodit и endopodit) конечностей (фиг. 45). У низших Р., например у веслоногих (Copepoda), не имеющих особых органов кровообращения, не существует и жабр; у таких Р. газовый обмен между кровью, находящейся в полости тела, и окружающей водой происходит на всех местах поверхности тела благодаря особенно тонким покровам тела. У высших Р. жабры имеют вид расщепленных или разветвленных пластинок или кустистых отростков (см. Конечности, фиг. 42—65), помещающихся на различных конечностях. Так, напр., у Amphipoda (фиг. 19—21) и Decapoda жабры помещаются на торакальных (грудных), а у равноногих (Isopoda, фиг. 81—82) и ротоногих (Stomatopoda, фиг. 25) на абдоминальных (брюшных) ножках. У десятиногих жабры прикрыты головогрудным щитком или его боковыми отростками, так что находятся в особой, так назыв. жаберной, полости (фиг. 84). Благодаря этому приспособлению жабры долгое время могут оставаться влажными, так что Р. могут пользоваться ими даже тогда, когда им приходится пребывать более или менее продолжительное время на воздухе. У некоторых представителей имеются даже особые приспособления для воздушного дыхания; так, у одного среднехвостого (Anomu r a) рака Birgus (см. Десятиногие раки), живущего на суше в норах, жаберная полость (фиг. 85) перегорожена загнутым внутрь краем головогрудного щитка на два отдела: верхний, довольно замкнутый, содержащий воздух и функционирующий наподобие легких отдел, и нижний, открытый — в котором находятся рудиментарные жабры.
Выделительные органы. Выделение последних продуктов обмена веществ совершается у Р. двояким путем: 1) при помощи накожных пор и 2) особых выделительных органов, т. е. раковинных желез или антеннальных желез. Раковинные железы встречаются у низших Р. (Entomostraca) и лишь у немногих Malacostraca, т. е. у некоторых Isopoda и Cumacea; они открываются наружу у основания второй пары челюстей и состоят из небольшого мешочка и извитого канальца, снабженного перед самым наружным отверстием небольшим резервуаром (мочевым пузырем). Антеннальные, или усиковые, железы встречаются у высших Р. (Malacoatraca) и имеют аналогичное строение, открываясь наружу у основания второй пары усиков. У Leptostraca встречаются раковинные и усиковые железы вместе, но первые являются рудиментарными, так что функционируют только усиковые железы. Выделительная функция этих желез была доказана Ковалевским лишь в недавнее время, что же касается их морфологического значения, то они соответствуют сегментальным органам червей.
Половые органы. Все Р., за очень немногими исключениями, раздельнополы. Гермафродитизм встречается лишь у некоторых сидячих и паразитических форм, а именно у сидячих и паразитических усоногих (Cirridia, ф. 14 и 16) и паразитических равноногих (Isopoda). Первые представляют весьма интересное биологическое явление, так как у некоторых из них, помимо обоеполых неделимых, встречаются еще пигмейные самцы, которые живут наподобие паразитов на теле (в складке мантии) гермафродитов. По наружному виду и внутренней организации самцы нисколько не походят на гермафродитных неделимов, а скорее на их личинки; они червеобразны, лишены ротового отверстия и имеют рудиментарный кишечник, усики и всего 4 пары недоразвитых конечностей. У других усоногих, имеющих пигмейных самцов, крупные и вполне развитые особи являются не обоеполыми, а самками, представляя, таким образом, пример полового диморфизма, столь распространенного среди Р. У паразитических усоногих и равноногих встречается протоандрический гермафродитизм, т. е. у обоеполых неделимых развиваются сначала мужские половые продукты и они функционируют как самцы, а затем, по атрофии первых и созревании женских половых продуктов, функционируют как самки. Рядом с этим у некоторых из них встречаются еще добавочные пигмейные самцы. Раздельнополые Р. отличаются обыкновенно и по наружному виду друг от друга: так, самцы (фиг. 2, 7, 9, 20, 21, 23, 24, 26—28 и 91) большей частью меньше и часто подвижнее самок, бывают снабжены обонятельными нитями на передних усиках и вооружены различными приспособлениями (видоизмененные конечности), служащими вспомогательными органами при совокуплении, т. е. для ловли, захвата и удерживания самки. У самок (фиг. 1, 3, 5—6, 8, 10—12, 18, 23а, 90), в свою очередь, встречаются особые приспособления, служащие для прикрепления и хранения оплодотворенных яиц, образующие у некоторых форм так назыв. выводковые полости, или камеры (фиг. 56), в которых из яиц развивается молодь. Мужские и женские половые органы Р. построены по одному типу и занимают одно и то же положение в теле. Они являются парными органами и только у некоторых Р. вследствие слияния между собой или появления промежуточного отдела становятся как бы непарными. В половых органах Р. (фиг. 86—91) отличают: 1) отделы, где приготовляются половые продукты, или у самцов семенники (testes), a y самок яичники (ovarii), которые в начале образования трудно отличимы друг от друга; 2) выводные протоки — у самцов семяпроводы (vasa deferentia) и яйцеводы (oviducti) y самок, 3) конечные отделы выводных протоков, отличимые анатомически и онтогенетически (эктодермального происхождения), а именно семяизвергательный канал (ductus ejaculatorius) y самцов и влагалище (vagina et vulva) у самок; у последних встречается еще у некоторых Р. семяприемник (receptaculum seminis); 4) органы совокупления, представляющие большей частью видоизмененные конечности. Будучи устроены по описанному типу, половые органы Р. варьируют только по форме и величине. Они помещаются обыкновенно на спинной стороне тела, между желудком и сердцем; половые отверстия находятся обыкновенно на брюшной стороне и только у некоторых Энтомострак (ветвистоусых и некоторых веслоногих) на спинной, причем у высших Р. (Malacostraca) в определенных сегментах, т. е. у самцов в последнем грудном (13) сегменте, а у самки в 3 грудном с конца (11); у низших Р. (Entomostraca) положение половых отверстий непостоянно у различных отрядов, но обыкновенно находится на границе переднего отдела и заднего, лишенного конечностей.
Размножение и развитие. P. размножаются исключительно половым путем, а бесполого размножения совершенно не встречается, точно так же, как нет и регенерации, а возможна только регенерация отдельных конечностей или частей их. В виде компенсации бесполого размножения у некоторых Р., а именно у листоногих (ветвистоусых и немногих жаберноногих) встречается партеногенез (см.). У этих форм самцы гораздо реже самок и появляются только периодически: так, у ветвистоусых лишь осенью. Тонкостенные летние яйца развиваются партеногенетически (без оплодотворения), и из них получаются самки, дающие несколько партеногенетических поколений. Осенью появляются, наконец, самцы, которые оплодотворяют более крупные, толстостенные, снабженные питательным материалом зимние яйца; последние перезимовывают, и весной из них развиваются вновь самки. Яйца громадного большинства Р. мезолецитальные, которые проделывают частичную, а именно поверхностную, сегментацию, и только у очень немногих Р. яйца имеют полное дробление. Лишь весьма немногие Р. проделывают прямое развитие, тогда как для большинства характерен своеобразный метаморфоз, заключающийся в постепенном превращении, сопровождающийся линькой, результатом которого являются весьма характерные свободноплавающие личинки. Так, для низших Р. (Entomostraca) характерна личинка науплиус (Nauplius, фиг. 92), которая покидает яйцо в виде овального, сплющенного в спинно-брюшном направлении организма с непарным глазом и лобным (фронтальным) органом чувств. На спинной стороне личинка прикрыта щитком, а на брюшной стороне несет три пары конечностей, из коих первая пара неразветвленная, дающая со временем 1-ю пару усиков (antennulae), вторая пара разветвленная, дающая 2-ю пару усиков (antennae), и третья пара, также разветвленная, дающая жвалы (mandibul a e). Кишечник состоит из трех отделов, и имеется нервная система. После линьки появляются челюсти (maxillae) и зачатки торакальных ножек, а задняя часть растет в длину и может образовать брюшко (abdomen). У паразитических Р. личинка прикрепляется к постороннему предмету и претерпевает регрессивный метаморфоз, выражающийся особенно рельефно у усоногих (см.). Для высших Р. (Malacostraca) характерна личинка зоэа (Zoea, фиг. 96). Из яйца, однако, вылупляется личинка науплиус (фиг. 93), которая проделывает затем ряд изменений или отдельных стадий — метанауплиус и протозоэа (фиг. 94 и 95) и которые были описаны в прежнее время как самостоятельные животные, пока более внимательное изучение их истории развития не показало, что мы имеем дело с личиночными формами. Изменения, происходящие с науплиусом, заключаются главным образом в появлении грудных конечностей, брюшка (abdomen) и конечностей на нем. В личинке Zoea (фиг. 96) различают головогрудь (cephalothorax) и брюшко (abdomen), парные глаза, кишечник, сердце и усиковые железы. Характерно, что десятиногие раки после зоэа проделывают мизидную стадию (фиг. 97), напоминающую по своей организации Р. Mysis, принадлежащего к расщепленогим (Schizopoda), из которых произошли десятиногие. Вообще личинки Р., их развитие, постепенное возникновение конечностей и других органов весьма характерны и в высшей степени поучительны и доказательны для эволюционной теории, как это блистательно изложено в вышедшем в 1864 г. сочинении Фр. Мюллер: "F ür Darwin".
Классификция. Р. подразделяются на две главные группы, или подкласса: низших Р., или Entomostraca, и высших Р., или Malacostraca, существенно отличающихся друг от друга как по внешнему виду, так и по внутренней организации, на что неоднократно указывалось в данной статье. I. Entomost r aca (фиг. 1—17), характеризующиеся непостоянным количеством сегментов, отсутствием брюшка или во всяком случае брюшных конечностей, присутствием раковинных железок и личинки науплиуса, делят на четыре отряда: 1) веслоногих или Copepoda (см., а также фиг. 1—6), которых подразделяют на Eucopepoda (свободноживущих или Gnathostoma, фиг. 1—2) и паразитических или Siphonostoma (фиг. 4—6) и Branchiura (фиг. 3). 2) Листоногих или Phyllopoda (см. соотв. ст., а также фиг. 7—11), которых подразделяют на жаберноногих или Brancniopoda (см. соотв. ст., а также фиг. 7—9) и ветвистоусых или Сlаdocera (см., а также фиг. 10—11). 3) Ракушковых или Ostracoda (см., а также фиг. 12) и 4) Усоногих или Cirripedia (см.), которых подразделяют на Thoracica, Abdominalia, Apoda и Rhizocephala, или корнеголовые (фиг. 13—17). II. Malacostraca (фиг. 18—28) характеризуются постоянным числом сегментов (20, из коих 13 передних образуют головогрудь, а 7 задних — брюшко), снабженных конечностями, присутствием челюстных ног и усиковой железы, а также положением половых отверстий в определенных (мужские в 11, женские в 13) сегментах и личинкой зоэа; этот подкласс делят на 3 главные группы: 1) Leptostraca (фиг. 18), представляющие как бы переход от Entomostraca к Malacostraca. 2) Члениcmогрудые раки, Arthrostraca (фиг. 19—23), у которых 7 грудных сегментов сливаются с головой в головогрудь, а глаза сидят на стебельчатых отростках. Группу Arthrostraca подразделяют на 2 отряда: a) амфиподы или Amphipoda (см., а также фиг. 19—21) и b) равноногие или Isopoda (см. выше, а также фиг. 22—23), которых подразделяют на Isopoda genuina sive Eoisopoda и Isopoda anomala sive Anisopoda. Группу Thoracostraca подразделяют на четыре отряда: a) расщепленогих или Schizopoda (см., а также фиг. 26—28), b) Cumacea (фиг. 24), c) Ротоногих или Stomatopoda (фиг. 25) и d) десятиногих раков или Decapoda (см.), которых подразделяют на длиннохвостых или Macrura, среднехвостых или Anomura и короткохвостых, или крабов, или Brachyura. Относительно характеристики отдельных отрядов, различных типичных представителей, а главное — образа жизни см. соответственные статьи.
Литература по Р., касающаяся отдельных отрядов, приведена в соответственных статьях, а поэтому ограничимся только списком сочинений, трактующих Р. в совокупности или некоторые общие вопросы по Р.: Milne-Edwards, "Histoire naturelle des Crustac é s" (П., 1834—1840, с атласом); H. Gerst äcker, "Bronn's Klassen u. Ordnungen des Thierreichs. Bd. V. Arthropoden" (I Abth. 1 Hälfte, 1866—79, 2 Hälft e, еще выходит выпусками); J. Dana, "Crustacea of M. S. exploring expedition under Captain Charles Wilkes" (Филадельфия, 1852, с атласом); С. Claus, "Untersuchungen zur Erforschung der genealogischen Grundlage des Crustaceensystems" (B., 1876); его же, "N eue Beiträ ge zur Morphologie der Crustaceen" ("Arbeit a. d. zool. Inst. zu Wien", т. VI, 1886); J. Boas, "Studien ü ber die Verwandschaftsbeziehungen der Malacostraken" ("Morphol. Jahrb.", т. VIII, 1883); С. Grobben, "Die Autennendr üsen der Crustaceen" ("Arb. aus d. zoolog. Inst. zu Wien", т. III, 1880); Fritz M üller, "Fü r Darwin" (Лпц., 1864); Th. Hutley, "The cray-fish. An introduction in to the study of Zoology" (Лпд., 1881, есть немецкий перев.); A. Lang, "Lehrbuch der vergleichenden Anatomie der wirbellosen Thiere" (Иена, 1894).
В. Шевяков.
Ракообразные (рыбов.) — составляют один из главнейших элементов пищи рыб, особенно в молодом их возрасте. Это обстоятельство дало мысль разводить их искусственно в бассейнах и прудах, предназначенных для помещения молоди разводимых рыб. На этом основано рыбоводное заведение города Люгрена (Lugrin) во Франции, в обиходе которого совершенно исключен всякий искусственный корм, заменяемый здесь "живой пищей". Ее составляют следующие Р.: бокоплав (Gammar us pulex), водяная блоха (Daphnia), Cypris, циклоп (Cyclops quadricornis) и мн. др. Благоприятные условия для быстрого размножения низших Р., как показывает опыт многих любителей аквариев и рыбоводов, создаются удобрением почвы органич. веществами, действием солнечных лучей на неглубокий слой воды и вымораживанием дна водоема зимой. Обыкновенно в качестве удобрения прибегают к птичьему помету, бросаемому в воду. Люгрен изобрел способ (составляющий его секрет) размножения Р. в любом количестве. По его расчету, в бассейне в 35 м длины, 23 м ширины и 0,40 м глубины можно в течение месяца получить от 300—350 килограммов Р. В такой бассейн можно пустить 20000 молоди форели, для которых требуется не менее 10 кг этой пищи в день. Форель при таком режиме растет очень быстро, достигая к 6 месяцам такого роста, что может идти на рынок. По мере поедания "живой пищи" в одном бассейне или пруде рыбешка переводится в другой, соседний, в котором также размножены Р. Этот же принцип с некоторым видоизменением проектирует ввести в прудовое хозяйство Воцелка (Wozelka). Сущность его проекта заключается в том, что по окраинам пруда, в котором кормится разводимая рыба, устраивается система мелких канавок, в которых дают свободно размножаться Р. Через решетчатые перегородки Р. свободно проскальзывают в соседние канавки для рыбы, которая переводится по мере использования из одного вместилища для разведения Р. в другое. Ср. Депп, "Размножение дафний" (Од., 1896); H. Бородин, "Рыбоводство" (стр. 58—64, 216); К. Wozelka, "Neues Fisc h zucht-Verfahren etc." (Прага, 1893).
|
Page was updated:Tuesday, 11-Sep-2012 18:16:17 MSK |