| [ начало ] | [ Т ] |
Ткани животные*
I. Эпителиальная Т. Плоский и призматический эпителий. Питание эпителиальной Т. Развитие эпителия. Железистый эпителий. — II. Соединительная Т. 1) собственно соединительная Т.: а) эмбриональная, b) ретикулярная, с) волокнистая, d) эластическая, е) жировая, f) пигментная Т. Развитие соединительной Т. 2) Хрящевая Т.: гиалиновый, сетчатый и волокнистый хрящ. Питание и развитие хрящевой Т. 3) Костная Т. — III. Мышечная Т.: а) гладкие, b) поперечно-полосатые мышцы, с) мышцы сердца. Кровеносные и лимфатические сосуды мышечной Т. Переход мышц в сухожилия. — IV. Нервная Т. Нервные клетки. Нервный отросток. Дендриты. Нервные волокна: мякотные и безмякотные. Развитие нервной Т. Невроглия. — V. Кровь и лимфа.
Простейшие, или Protozoa, в течение всей своей жизни состоят только из одной клетки, которая несет все функции, необходимые для существования каждого такого организма. Следующие за ними более сложные организмы, или Metazoa, лишь в известной стадии своей жизни представляются в виде одной клетки — женского яичка, которая, соединившись с другою, семенною клеткою — в одно целое, претерпевает затем ряд весьма интересных процессов, влекущих за собою в конце концов образование того или иного сложного организма. Процессы эти состоят в многократном делении яйца, в распределении вновь возникших клеток на отдельные пласты — зародышевые листки — и в постепенной, возрастающей по мере усложнения, организации, дифференцировке или специализации труда между отдельными клетками. Клетки, имеющие приблизительно одинаковое строение, форму и химический состав, слагаются в особенные клеточные группы; одновременно с этим клетками вырабатываются особенные вещества, которые располагаются в большем или меньшем количестве в промежутках между клетками и носят назв. межклеточных, или основных, веществ. Такого рода группы одинаково дифференцированных клеток с их производными составляют то, что мы наз. "простыми Т.". В свою очередь простые Т., вступая в известные отношения между собою, образуют "сложные Т.", или "органы". К простым Т. относятся: 1) эпителиальная Т., 2) поддерживающая — соединительная Т., 3) мышечная и 4) нервная Т. Распределяя группы клеток на Т., руководствуются не одною какою-либо особенностью той или другой клеточной группы, а принимают обыкновенно во внимание много различных свойств, которые характеризуют данную Т. и дают возможность отличить ее от других Т. К числу признаков, принимаемых в основу деления клеточных групп, принадлежат: строение и химический состав каждой группы, ее развитие и физиологическая функция. Пользуясь указанными признаками, всегда можно отличить элементы одной какой-либо простой Т. от элементов других Т.
I. Эпителиальная ткань (табл. I, эндотелий и эпителий), как показывает самое название (έπι - τέλλω — настилаю, выстилаю) образует наружные покровы тела и выстилает собою полости различных органов. Кроме того, из нее построены различные придатки кожи — волосы, иглы, перья, а равно ногти, копыта, рога и пр. Далее, эпителиальная Т. входит в состав многих органов, известных под назв. "желез", как, напр., слюнные железы, печень, почки и т. п. Наконец, к эпителиальной Т. относятся особенные клетки, служащие для воспринятия известных впечатлений из внешнего мира, которые они затем передают нервам. Такого рода эпителий назыв. "нервным эпителием" ("невроэпителием") и встречается в органах чувств, напр. в органе вкуса и др. Характерные особенности эпителиальной Т. заключаются в следующем: а) она состоит почти исключительно из клеток, связанных между собою лишь весьма незначительным количеством межклеточного вещества; b) в ней нет кровеносных сосудов (исключение составляют — слизистая оболочка нёба у амфибий, орган слуха, эпителий слизистой оболочки reg. olfactoriae); с) эпителиальная Т. всегда помещается на соединительнотканной подстилке. Клетки эпителиальной Т. имеют самую разнообразную форму, которая до известной степени стоит в зависимости от характера и функции данного органа. Однако, несмотря на разнообразие форм эпителиальных клеток, между ними все-таки легко можно отличить две основных формы: клетки в виде более или менее толстых многоугольных пластинок и в форме коротких или длинных призм (цилиндров) и пирамид (конусов). Принимая во внимание указанные основные формы, всю эпителиальную Т. разделяют на две группы: на так назыв. плоский (мостовидный) и на призматический (цилиндрический) эпителий. Обыкновенно клетки плоского и призматического эпителия, располагаясь одна возле другой, образуют один или много клеточных рядов (слоев). В первом случае эпителий получает назв. однорядного (однослойного) плоского или призматического — цилиндрического — эпителия; во втором случае он назыв. многорядным (многослойным), плоским или призматическим эпителием. Нередко от свободного конца клеток однорядного и многорядного призматического эпителия отходят особенные придатки, имеющие форму более или менее длинных и тонких нитей (волосков или ресниц), которые при жизни находятся в постоянном движении. Такой эпителий назыв. мерцательным (ресничным) призматическим эпителием.
Плоский эпителий (фиг. 1—4). Особенности этого эпителия заключаются в том, что образующие его клетки имеют форму то более, то менее тонких, большей частью многоугольных пластинок с неровными краями. Если клетки располагаются лишь в один ряд, все они имеют приблизительно одинаковую толщину, наоборот, если они распределяются друг над другом в несколько рядов — толщина их в разных рядах бывает неодинакова. Обыкновенно клетки самого глубокого ряда (слоя), прилегающие непосредственно к соединительнотканной подстилке, имеют довольно значительную высоту и кажутся в виде кубической, призматической или булавовидной формы образований (фиг. 3). Клетки следующих за ним нескольких рядов представляются в форме многогранных, сравнительно толстых пластинок, на поверхностях которых замечается ряд более или менее глубоких вдавлений и гребешков. Затем толщина клеток следующих рядов постепенно уменьшается по мере приближения данного ряда к свободной поверхности эпителиального покрова (фиг. 3 и 4). Наконец, клетки самых поверхностных рядов, за немногими лишь исключениями (напр., клетки эпителия мочевого пузыря), имеют форму тончайших пластинок (фиг. 3). Протоплазма эпителиальных клеток составлена из тончайших ниточек — фибриллей, переплетающихся между собою в разных направлениях и в то же время связанных друг с другом; промежутки между фибриллями заняты межфибриллярным веществом (фиг. 2). В каждой клетке помещается одно, иногда два больших, круглой или овальной формы ядра, с одним или несколькими ядрышками (фиг. 1—4). В клетках, имеющих вид тонких пластинок, ядра кажутся более или менее сплющенными или даже могут совсем отсутствовать. Но нередко рядом с одно- и двуядерными клетками встречаются клетки со многими ядрами. Величина эпителиальных клеток так же разнообразна, как и форма их. Большинство эпителиальных клеток не имеет оболочки. В протоплазме клеток как однорядного, так и многорядного эпителия — нередко встречаются различного рода включения: а) зернышки желтого, бурого или черного пигмента (клетки однорядного плоского эпителия сетчатки, в глубоких рядах многослойного эпителия кожи, в особенности у цветных рас, напр. негров, и у животных); b) зернышки особенного альбуминоидного вещества — элеидина, не растворимые в воде, эфире, алкоголе и пр. (в клетках многослойного мостовидного эпителия кожи). Из означенных зернышек, по мнению некоторых исследователей, образуется потом альбуминоидное тело кератин — роговое вещество, заключающееся в клетках поверхностных рядов эпителия кожи. Присутствие этого вещества в клетках делает их плотными, упругими, способными противостоять действию кислот, щелочей и пр. и превращает их в конце концов в роговые пластинки (чешуйки). Между всеми клетками эпителия помещается весьма незначительное количество особого, полужидкой консистенции, межклеточного (склеивающего) вещества, при помощи которого клетки теснее связываются друг с другом. Межклеточное вещество обыкновенно выступает ясно лишь после обработки эпителия известными реактивами, к которым относятся слабые растворы азотнокислого серебра и метиленовой сини. От действия указанных реактивов границы между клетками обозначаются в виде черных (фиг. 1) или синих линий. Кроме того, от клеток еще отходят весьма нежные и тонкие отростки, состоящие из нитчатого вещества протоплазмы. Они, наподобие мостиков, переходят через межклеточное вещество, направляясь из тела одной клетки в тело других, окружающих ее клеток и имеют вид тончайших ниточек (фиг. 2 и 3). Означенные ниточки известны под названием "межклеточных" или "протоплазматических мостиков" и обыкновенно при надлежащей обработке эпителия растворами азотнокислого серебра или метиленовой сини остаются неокрашенными, вследствие чего границы между клетками кажутся в виде прерывистых линий. Между эпителиальными клетками находятся как постоянные, так равно и временные элементы, которые, однако, не имеют ничего общего с эпителием; к первым относятся нервы, ко вторым — лейкоциты. Что касается нервов, то пока присутствие их доказано только в многорядном мостовидном эпителии, причем они имеют вид тончайших, постепенно делящихся ниточек и представляют собою конечные разветвления чувствительных нервов. Означенные разветвления нередко, как, напр., в эпителии роговицы, достигают почти самых поверхностных рядов эпителиальных клеток и образуют вокруг последних более или менее густое сплетение. Лейкоциты попадают в эпителий из различного рода полостей, наполненных лимфою, серозною жидкостью или кровью (из серозных полостей, лимф. и кровеносных сосудов) в виде так называемых "блуждающих", или "странствующих", клеток или же из подлежащей соединительной Т. В плоском однорядном эпителии лейкоциты обыкновенно проникают через вещество, склеивающее клетки, причем на местах прохождения их остаются в течение известного времени небольшие отверстия — устьица (stigmata или stomata). В многорядном мостовидном эпителии лейкоциты располагаются между клетками и, передвигаясь, достигают свободной поверхности эпителиального покрова. Таким образом они выселяются в полости различного рода органов (в полость рта, глотки, кишечного канала и др.). В тех органах, где в подэпителиальной соединительной Т. находятся так называемые лимфатические узелки, или фолликулы, выселение лейкоцитов в эпителий достигает колоссальных размеров. Они нередко занимают почти все промежутки между эпителиальными клетками, маскируют эти последние и даже влекут за собою отпадение целых эпителиальных пластов. Излюбленными местами выселения лейкоцитов являются: миндалевидные (лимфат.) узлы, слизистая оболочка корня языка, свода глотки и место перехода тонких кишок в толстые и пр., где имеются скопления лимфатических узелков. Однорядный плоский эпителий выстилает внутреннюю поверхность стенки кровеносных и лимфатических сосудов, поверхности серозных оболочек, т. е. плевры, сердечной сумки, брюшины, а равно полости, наполненные жидкостью (лимфою), как, напр., суставные полости, полость передней камеры глаза и др. Кроме того, означенный эпителий покрывает внутреннюю поверхность легочных пузырьков, мелких бронхов и выводные протоки многих желез; в двух последних случаях он является уже в виде кубического эпителия. Многорядный мостовидный эпителий покрывает всю поверхность кожи человека и др. позвоночных, слизистые оболочки полости рта, глотки и пищевода, переднюю поверхность роговицы, слизистую оболочку мочеиспускательного канала, мочевого пузыря, мочеточников и почечных лоханок, слизистую оболочку влагалища и пр. Эпителий, выстилающий внутреннюю поверхность указанных мочевых органов, называется "переходным эпителием", так как в состав его входит лишь немного (4—5, иногда больше) рядов клеток, причем клетки самого поверхностного ряда имеют вид больших и довольно толстых многоугольных пластинок; в них в большинстве случаев находится несколько крупных ядер.
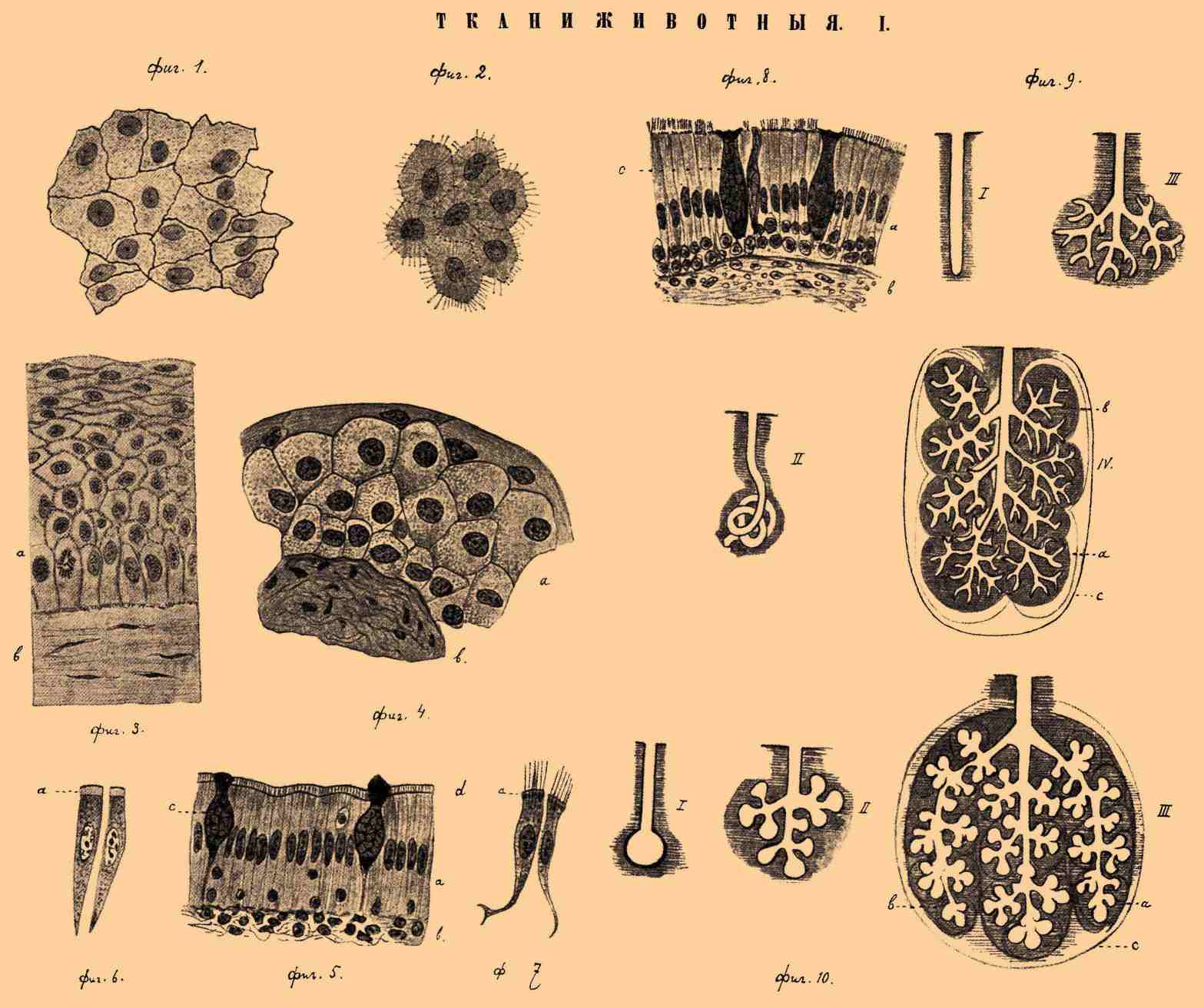
ТКАНИ ЖИВОТНЫЕ I.
Фиг. 1. Плоский однорядный эпителий. Сердечная сумка кошки. Увелич. в 200 раз. Фиг. 2. Плоский однорядный эпителий, покрывающий заднюю поверхность роговой оболочки глаза собаки. Видны нити и межнитчатое вещество протоплазмы клеток, межклеточное вещество и протоплазматические мостики. Увелич. в 500 раз. Фиг. 3. Часть разреза роговой оболочки глаза человека. a — эпителий; b — подлежащая соединительная ткань. Увелич. в 500 раз. Фиг. 4. Часть разреза мочевого пузыря мыши. а — многорядный плоский эпителий; b — подлежащая соединительная ткань. Увелич. в 500 раз. Фиг. 5. Часть разреза тонкой кишки кошки. a — однорядный призматический (цилиндрический) эпителий; b — подлежащая соединительная ткань; с — бокаловидные слизистые клетки; d — кутикулярный ободок. Увелич. в 500 раз. Фиг. 6. Две изолированные клетки однорядного призм. эпителия из тонкой кишки кошки, а — кутикулярный ободок. Увел. в 500 раз. Фиг. 7. Две изолированные клетки из многорядного мерцательного эпителия дыхательного горла собаки, a — утолщенные (базальные) концы мерцательных волосков. Увелич. в 500 раз. Фиг. 8. Часть разреза дыхательного горла кошки. а — многорядный призматический мерцательный эпителий; b — подлежащая соединительная ткань. Увелич. в 500 раз. Фиг. 9. Трубчатые железы. I — простая трубчатая железа; II — простая трубч. железа, конец которой изогнут наподобие клубочка; III — простая ветвящаяся трубчатая железа; IV — сложная ветвящаяся трубчатая железа. a — доли; b — дольки; с — соединительнотканная оболочка. Схематические рисунки. Фиг. 10. Альвеолярные (пузырчатые) железы. I — простая альвеолярная железа; II — ветвящаяся альвеолярная железа; III — сложная ветвящаяся альвеолярная железа, a — доли; b — дольки; с — соединительнотканная оболочка. Схематические рисунки.
Призматический или цилиндрический эпителий (фиг. 5—8) характеризуется тем, что в состав его входят более или менее высокие клетки призматической и пирамидальной формы, почему данное ему и общепринятое название "цилиндрический эпителий" не отвечает действительности. В многорядном призматическом эпителии обыкновенно в промежутках между суженными нижними (внутренними) концами клеток призматической или пирамидальной формы помещаются в один или несколько рядов клетки полиэдрической, веретенообразной и неправильной формы, вследствие чего эпителий кажется составленным из нескольких рядов клеток. Нередко верхний (свободный) конец призматических клеток бывает покрыт кутикулой, напр. в однорядном призматическом эпителии кишок (фиг. 5 и 6), или же от него отходит множество тонких нитевидных придатков — мерцательных волосков или ресниц (фиг. 7 и 8), как это имеет место в однорядном эпителии матки, мелких бронхах и пр. и в многорядном эпителии, выстилающем слизистую оболочку гортани, дыхательного горла и др. Подобного рода эпителий носит название призматического однорядного или многорядного мерцательного — ресничного — эпителия. Среди клеток однорядного и многорядного призматического эпителия постоянно встречаются особенные клетки, в протоплазме которых помещаются капельки слизи (муцина), отчего эти клетки получили название "бокаловидных слизистых клеток" (фиг. 5 и 8с). Бокаловидными их называют потому, что от содержащего в них секрета — слизи — клетки кажутся вздутыми наподобие рюмки, бокала или бочонка; только нижний конец клеточного тела, не заключающий в себе слизи, представляется то более, то менее суженным, и в нем обыкновенно располагается несколько сдавленное клеточное ядро. Бокаловидные клетки происходят из клеток призматических или пирамидальных эпителия путем постепенного превращения известной части их протоплазмы в слизь. Протоплазма описываемого эпителия также представляется дифференцированною на нити — фибрилли — и межнитчатое вещество, причем, по мнению некоторых исследователей, нити плазмы переходят, как и в плоском эпителии, из одной клетки в другие в виде протоплазматических мостиков. Между клетками помещается небольшое количество мягкого межклеточного вещества, которое часто в верхних отделах эпителиального покрова, вблизи его свободной поверхности, становится более плотным и образует так назыв. "замыкающие полоски". В среднем или в более толстом верхнем отделе каждой клетки или часто ближе к нижнему ее концу располагаются одно, а иногда и два овальной формы и довольно больших ядра со всеми своими составными частями. Между клетками призматического эпителия имеются тонкие нервные нити, оплетающие клетки, а равно часто попадаются и лейкоциты, которые иногда даже вползают внутрь (в протоплазму) клеток. Однорядный призматический эпителий покрывает слизистую оболочку всего желудочно-кишечного канала, желчного пузыря, а равно более толстые из выводных протоков многих желез, напр. собирательные канальцы почек, выводные трубки слюнных желез и др. Многорядный призматический эпителий встречается лишь в очень немногих органах, а именно — покрывает собою свод конъюнктивы (слизистой оболочки век) и слизистую оболочку мочеиспускательного канала, где он находится островками среди клеток многорядного плоского эпителия. Однорядный и многорядный мерцательный призматический эпителии выстилают: первый слизистую оболочку мелких бронхов, матки, яйцеводов и пр., второй — слизистую оболочку респираторного отдела полости носа, гортани, дыхательного горла и крупных бронхов, каналы придатка яичка (epidydimis) и выносящего семя протока (vas deferens) и пр.
Питание эпителиальной ткани. Эпителий не имеет кровеносных сосудов, за немногими лишь исключениями, а потому исследователи уже давно старались выяснить, каким образом совершается питание этой ткани. Наблюдения, произведенные над плоским и цилиндрическим эпителием, показали, что питание его происходит путем межклеточных промежутков, наполненных полужидким веществом. Протоплазматические мостики, пронизывающие означенные промежутки, служат не столько для более прочной связи между клетками, сколько для того, чтобы препятствовать их тесному сближению и постоянно удерживать клетки на известном расстоянии друг от друга. Благодаря означенному приспособлению, межклеточные промежутки остаются образованиями постоянными и сообщаются непосредственно с соконосными канальцами и щелями, а при помощи их и с лимфат. сосудами подлежащей соединительной Т. В многослойном мостовидном эпителии межклеточные промежутки ясно выражены лишь в глубоких рядах клеток, между тем как в поверхностных рядах они выступают неясно и передвижения по ним питательной жидкости, вероятно, не происходит. Постоянно, в течение всей жизни животного, многие из старых эпителиальных клеток умирают и замещаются новыми элементами, которые в многорядном плоском эпителии происходят путем митотического деления некоторых клеток самого глубокого ряда. Последний вследствие этого называется "ростковым слоем".
Развитие эпителия происходит из всех трех зародышевых листков: плоский многорядный эпителий наружных покровов тела, полости рта и пр. развивается из эктодермы; однорядный призматический эпителий желудочно-кишечного канала, желчного пузыря, выводных протоков многих желез, и т. д. получает начало из энтодермы, а однорядный плоский эпителий, выстилающий внутреннюю поверхность стенок кровеносных и лимфатич. сосудов и пр., развивается из мезодермы.
Железистый эпителий представляет собою особенный вид эпителиальной ткани. Клетки этого эпителия обладают способностью вырабатывать известного рода вещества, необходимые для жизни организма, так назыв. секреты, или же выделять различного рода продукты обмена веществ, ненужные и вредные для организма, которые называют экскретами. Железистые клетки то располагаются отдельно среди клеток того или другого эпителия, как, напр., бокаловидные слизистые клетки, то собираются в целые комплексы и в таком случае образуют более или менее сложные органы — слюнные железы, печень, почки и др. В первом случае клетки железистого эпителия получают название одноклеточных желез, во втором, когда ими составляются органы, их назыв. многоклеточными железами или просто железами. В одноклеточных железах каждая отдельная клетка функционирует независимо от других подобных же клеток, причем приготовляемое ею вещество непосредственно выбрасывается на поверхность тела животного или в полость какого-либо органа. Подобные клетки, как, напр., упомянутые выше бокаловидные клетки, помещаются между клетками призматического однорядного как мерцательного, так и немерцательного эпителия, не углубляясь в подлежащую ткань (фиг. 4 и 8); но иногда они выходят за границы эпителия и более или менее глубоко внедряются в находящуюся под эпителием соединительную ткань, напр. у многих низших животных. В последнем случае железистые клетки приобретают значительную величину и та часть их, которая лежит в соединительной ткани, кажется вздутою, а остальная часть, расположенная между эпителиальными клетками, вытягивается наподобие шейки; в общем описываемые клетки получают вид колбы с длинной шейкой. Бокаловидные клетки образуются, бесспорно, из обыкновенных покровных клеток призматического и мерцательного эпителия, покрывающего, напр., полость желудочно-кишечного канала, гортань, дыхательное горло и пр. Превращение протоплазмы этих клеток в слизь (муцин) обыкновенно начинается с верхнего (обращенного в сторону полости данного органа) конца клетки, в котором сначала появляется несколько небольших капелек секрета — секреторных вакуолей. Постепенно количество этих капелек увеличивается, многие из них сливаются в капли больших размеров, а вместе с этим тело клетки вздувается, становится округленным, причем капельки секрета, увеличиваясь в количестве и в размерах, оттесняют часть протоплазмы с ядром к нижнему концу клетки. По мере накопления в клетке секрета она становится светлее, за исключением лишь того ее отдела, в котором заключается ядро с не измененной еще протоплазмой. Капли секрета, занимающие в известной стадии деятельности клетки большую ее часть, отделяются друг от друга лишь тонкими прослойками из неизмененной протоплазмы, а по всей периферии клетки последняя, уплотняясь, образует род оболочки. Клеточное ядро сдавливается в большей или меньшей степени, причем если клетка имела в начале кутикулярный ободок или мерцательные волоски, то обыкновенно эти образования постепенно утрачиваются клеткою. Когда секрета накопится в клетке достаточное количество, тогда она принимает форму почки или бокала с более или менее длинною ножкой и наконец у верхнего своего конца лопается, а секрет в виде комочков слизи выделяется на свободную поверхность органа. Вслед за выделением секрета железистая клетка уменьшается в объеме, сдавливается окружающими клетками и становятся едва заметной. По истечении известного времени освободившаяся от секрета клетка вновь принимает прежний свой вид, восстановляется, вероятно, на счет остатка неизменившейся протоплазмы. Таким образом, одна и та же клетка много раз может приготовлять и выделять секрет и затем лишь погибает, уступая место новым клеткам. Что касается многоклеточных желез, то в них железистые клетки собираются в группы, причем каждая клетка функционирует одновременно с другими окружающими ее клетками. Обыкновенно железистые клетки располагаются одна возле другой в один ряд или же в несколько рядов и представляются то в виде многоугольных пластинок или кубов, то в форме призм и пирамид (конусов). Располагаясь друг возле друга, железистые клетки ограничивают собою полость (канал), в которую поступают вырабатываемые или выделяемые клетками вещества, и в то же время составляют стенку этой полости, т. е. стенку железы. Кнаружи от эпителиального покрова железы располагается тонкая, бесструктурная соединительнотканная оболочка, так назыв. собственная оболочка — membrana propria; она отделяет железистый эпителий от окружающих его частей — соединительной Т., кровеносных сосудов и пр. Полость, стенку которой составляет железистый эпителий, имеет вид более или менее длинного канала, или же конец этого канала представляется расширенным наподобие пузырька или мешочка, вследствие чего и все железы разделяются на так назыв. трубчатые (фиг. 9) и альвеолярные (фиг. 10) (пузырчатые — от слова alveolus — пузырек) железы. В свою очередь трубчатые железы являются или в виде простых неветвящихся трубочек, напр. железы дна желудка, кишечные или Либеркюновы железы и др., и в таком случае получают название простых трубчатых желез (фиг. 9, I). Обыкновенно один конец каждой такой трубки открывается на свободную поверхность какого-либо органа, а другой оканчивается слепо; иногда нижний конец трубочки кажется свернутым наподобие клубочка, напр., в потовых железах (фиг. 9, II). Или же подобная трубочка разделяется на несколько в свою очередь делящихся трубочек — тогда железы получают название простых трубчатых ветвящихся желез (фиг. 9, III), к которым принадлежат, напр., железы выхода желудка (пилорные), Бруннеровы железы и др. В этих железах та более толстая трубка, в которую собираются все остальные трубочки, назыв. выводным ее протоком, а остальная часть трубочек представляет собою тело железы. В большинстве случаев лишь эпителий трубочек, составляющих железистое тело, приготовляет секрет или играет роль выделительного органа, между тем как эпителий выводного протока имеет значение лишь покровного эпителия. В том случае, когда выводной проток ветвящейся трубчатой железы многократно делится, а трубочки, составляющие тело железы, в свою очередь распадаются на множество изогнутых трубочек — железа получает название сложной ветвящейся трубчатой железы (ф. 9, IV). Благодаря многократному делению трубочек подобные железы значительно увеличиваются в объеме и нередко получают вид крупных органов. Обыкновенно снаружи такие железы окружаются более или менее толстою соединительнотканною оболочкою (ф. 9, IV с), от которой отделяются прослойки в промежутки между системами трубочек и разделяют железу на участки — доли (lobi; фиг. 9, IV a). От этих прослоек в свою очередь отходят более тонкие прослойки в глубину долей, вследствие чего каждая доля разбивается на известное количество более мелких участков — долек (lobuli; фиг. 9, IV b). К сложным ветвящимся трубчатым железам относятся: поджелудочная железа (pancreas); слезные железы, почки и др. Иногда трубочки, образующие тело этих желез, соединяются между собою и, составляя целую сеть трубочек, получают название сетевидных желез, как, напр., печень. Что касается альвеолярных (пузырчатых) желез, то они также разделяются на простые альвеолярные железы (фиг. 10, I), если неделящиеся трубочки оканчиваются вздутиями наподобие пузырьков или мешочков; к ним причисляют кожные железы рептилий и амфибий, мелкие сальные железы и др. Под названием простых ветвящихся альвеолярных желез (фиг. 10, II), как, напр., крупные сальные железы, понимают такие железы, в которых концы делящихся трубочек расширяются в пузырьки. Наконец, сложные ветвящиеся альвеолярные железы (фиг. 10, III) вполне аналогичны таким же трубчатым железам и отличаются от них лишь тем, что трубки их оканчиваются различной величины и формы расширениями. К этим железам причисляют в последнее время слюнные железы полости рта, затем молочную железу и легкие. Во всех сложных ветвящихся железах, как видно из вышеизложенного, всегда имеется один толстый, иногда довольно длинный выводной проток, который помещается вне массы железы, и много мелких протоков, на которые он непосредственно подразделяется; мелкие протоки помещаются в теле самой железы и идут в нем по прослойкам соединительной Т., разделяющей железу на доли и дольки. Помимо всех перечисленных желез, имеются еще железы без выводных протоков; к подобным железам принадлежат все так назыв. кровяные железы, как, напр., щитовидная и надпочечная железа и др. Вещества, вырабатываемые клетками этих желез, попадают непосредственно через кровеносные и лимфатические сосуды в кровь и служат как бы для нейтрализации известных вредных продуктов обмена веществ, которые поступают из Т. и органов в кровеносную систему. Вырезывание этих желез влечет за собою смерть организма или сильные расстройства его нормальной деятельности. Продукты, приготовляемые или выделяемые железами, представляются или в виде более или менее вязкой, тягучей и содержащей слизь жидкости, или в форме водянистой жидкости, заключающей в себе растворы различных белковых веществ, или же, наконец, имеют вид густой жировой массы. Принимая во внимание характер секретов и экскретов, все железы можно бы разделить на три группы: на слизистые, серозные, или белковые, и сальные железы. Часто клетки одной и той же железы приготовляют как слизь, так и богатую белком жидкость и в таком случае получают название смешанных желез. Различные вещества, приготовляемые железами, выделяются в просвет, ограниченный железистыми клетками, и оттуда через выводной проток уносятся вон из желез, причем самые клетки не погибают, как, напр., мы это видим в слюнных железах, печени и др. Во многих из таких желез, напр. во всех серозных железах, секрет, прежде чем попасть в просвет железистых трубок или пузырьков, поступает предварительно в тонкие канальцы, окружающие клетки или находящиеся внутри клеток. Трубочки эти носят название секреторных капилляров (подробности см. Слюнные железы). Рядом с этим имеются и такие железы, секрет которых происходит путем разрушения частей самых железистых клеток, как, напр., в молочной железе, или даже целых клеток, как мы это видим в сальных железах. Все вообще железы очень богаты кровеносными, лимфатическими сосудами и нервами, которые помещаются в сложных ветвящихся железах в соединительнотканных прослойках. Мелкие разветвления кровеносных сосудов — капилляры — прилегают обыкновенно непосредственно к стенкам железистых трубок и пузырьков, отделяясь от железистых клеток лишь тонкою собственною оболочкою — membr. propria — желез. Конечные нервные разветвления в большинстве желез проникают в промежутки между железистыми клетками (подр. см. Сальные и Слюнные железы). Железы развиваются путем врастания — впячивания — эпителиальных клеток, покрывающих тот орган, на поверхность которого открываются выводные протоки этих желез.
II. Соединительная, или поддерживающая, Т., как показывает самое название, служит для соединения и для поддержки различных органов и частей тела животного организма. Нет такой Т. и органа, за немногими лишь исключениями, в состав которых не входило бы большее или меньшее количество поддерживающей Т., почему она и включается в число самых распространенных простых Т. организма человека и животных. Отличительным признаком этой Т. служит то, что в состав ее входит большое сравнительно количество так называемого основного вещества и лишь незначительное количество клеток. Изучение развития поддерживающей Т. показывает, что основное (межклеточное) вещество происходит из клеток и, постепенно дифференцируясь, приобретает известные характерные особенности. Оно или превращается в пучки тончайших нитей (фибриллей), или становится, по-видимому, однородным, довольно плотным и упругим, или же, наконец, пропитывается солями извести и вследствие этого приобретает значительную твердость. Одновременно с дифференцировкой основного вещества изменяются несколько химический его состав и форма соединительнотканных клеток. Принимая во внимание строение и отчасти химический состав основного вещества, отличают три рода соединительной Т.: 1) собственно соединительную (волокнистую) Т., 2) хрящевую и 3) костную Т. Несмотря на кажущееся резкое различие между означенными тремя формами соединительной Т., они, однако, по своему строению и химическому составу имеют очень много общего и происходят из среднего зародышевого листка (мезодермы).
1) Собственно соединительная Т. характеризуется тем, что основное вещество ее представляется мягким, имеет ясно выраженное волокнистое строение и при варении дает животный клей (глутин) или содержит муцин или эластин. Что касается соединительнотканных клеток, то большинство их имеют вид тонких отростчатых пластинок. Элементы собственно соединительной ткани — волокна и клетки — могут до известной степени изменяться сообразно с характером того органа, основу которого они составляют. Так, напр., в одном случае волокна идут в каком-либо одном направлении, в другом — в разных направлениях или даже совсем отсутствуют; иногда клеток в означенной Т. очень много, иногда, наоборот, мало или же в них находятся различные включения (жир, пигмент и пр.). Принимая в соображение те или другие особенности описываемой Т., ее для удобства самого изучения разбивают на несколько групп или видов, к которым относят: а) эмбриональную соединительную Т., b) ретикулярную Т., с) волокнистую (фибриллярную) соединительную Т., d) эластическую, е) жировую и f) пигментную Т. а) Эмбриональная (слизистая) ткань (табл. II, фиг. 1) находится исключительно у зародышей в тех местах тела, где впоследствии образуется тот или другой род поддерживающей Т. (напр., волокнистая или жировая Т.); она входит также в состав так назыв. Вартоновой студени пуповины. Основное вещество этой Т. у очень молодых зародышей кажется совершенно однородным, бесструктурным и содержит большее или меньшее количество слизи (муцина), почему некоторые исследователи называют эту Т. "слизистою тканью". У более взрослых зародышей в основном веществе эмбриональной Т. уже появляются тонкие волоконца, составленные из тончайших ниточек, которые склеены между собою ничтожным количеством особенного вещества. В основном веществе располагается множество плоских отростчатых клеток с довольно большими круглыми или овальными ядрами. Обыкновенно отростки клеток, многократно делясь, переплетаются и соединяются друг с другом, вследствие чего возникает род более или менее густой сети, промежутки которой заняты основным веществом. В ядрах клеток этой Т. часто встречаются митотические фигуры, указывающие на усиленное размножение клеток. b) Ретикулярная (аденоидная) Т. (табл. II, фиг. 2) составляет остов известных, преимущественно кровообразовательных, органов, к которым относятся: селезенка, костный мозг, лимфатические узлы (железы), солитарные узелки (фолликулы). Всюду, где только встречается эта Т., основное вещество ее состоит из очень тонких волоконец, которые соединяются между собою и образуют более или менее узкопетлистую сеть — reticulum. Каждое такое волоконце в свою очередь является составленным из тончайших ниточек — фибриллей, которые склеиваются между собою незначительным количеством особенного склеивающего (межфибриллярного) вещества. Таким образом, волоконца на самом деле суть пучки тончайших соединительнотканных фибриллей. От продолжительного действия кислот и едких щелочей пучки фибриллей набухают и растворяются, но при варении они не дают животного клея (глутина). На местах соединения пучков, в так наз. узловых точках сети, обыкновенно возникают угловатой формы расширения, к поверхности которых прилегают тела плоских, звездчатой формы клеток ретикулярной Т. В клетках помещаются довольно большие, круглой формы ядра, а самые клетки и их отростки, прилегая к пучкам фибриллей, покрывают их. В петлях сети ретикулярной Т. располагаются различного рода клеточные элементы, входящие в состав кровообразовательных органов. К таким элементам относятся: лейкоциты — лимфоциты (в лимфатических узлах); лейкоциты, красные кровяные клетки с ядрами (молодые), многоядерные гигантские клетки и др. (в костном мозгу и селезенке). Ввиду того, что вся сетка ретикулярной Т. набита указанными элементами, ее можно видеть лишь при известных условиях, а именно, когда эти элементы будут удалены из Т., напр. путем осторожного выбивания тонких разрезов органов кисточкой. с)
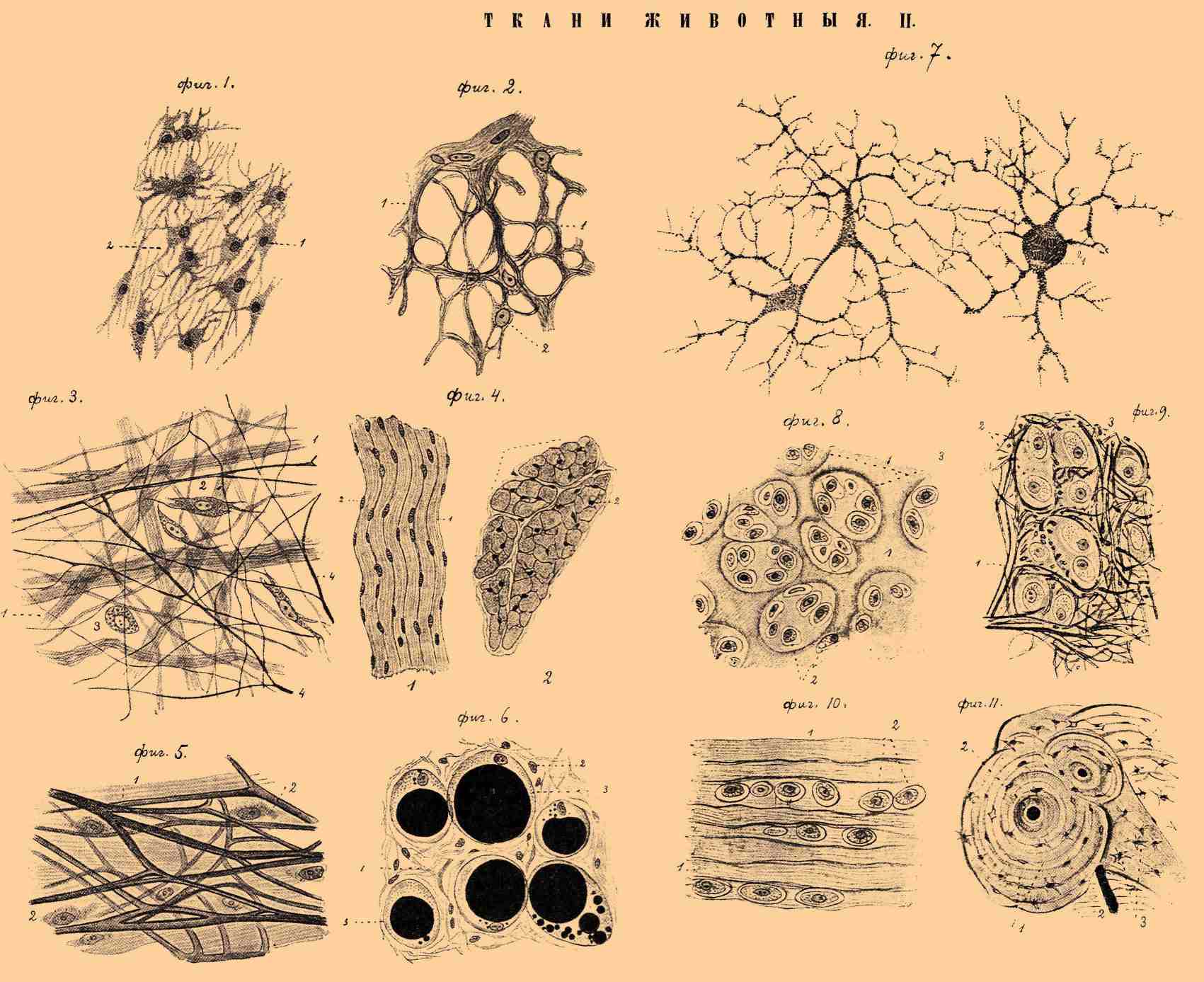
ТКАНИ ЖИВОТНЫЕ II.
Фиг. 1. Эмбриональная ткань из околоплодной оболочки зародыша свиньи. 1 — клетки; 2 — ядра. Увелич. в 240 раз. Фиг. 2. Ретикулярная ткань из лимфатич. узла собаки. 1 — пучки фибриллей; 2 — ядра клеток. Увелич. в 535 раз. Фиг. 3. Рыхлая волокнистая соединительная ткань из подкожной жирной клетчатки крысы. 1 — пучки фибриллей; 2 — плоские клетки, 3 — тучная клетка (Эрлиха); 4 — эластические волокна Увелич. в 535 раз. Фиг. 4. Плотная соединительная ткань. I — часть продольного и поперечного (II) разрезов сухожилия теленка. 1 — пучки фибриллей; 2 — ядра клеток. Увелич. в 240 раз. Фиг. 5. Эластическая ткань из выйной связки теленка. 1 — сеть эластических волокон; 2 — клетки волокнистой соединительной ткани. Увелич. в 535 раз. Фиг. 6. Жировая ткань из подкожной жирной клетчатки человека. 1 — пучки фибриллей; 2 — ядра клеток; 3 — жировые клетки с включенными в них каплями жира. Увелич. в 240 раз. Фиг. 7. Пигментная ткань из кожи личинки саламандры. 1 — пигментная клетка с делящимся ядром. Увелич. в 240 раз. Фиг. 8. Хрящевая ткань. Часть разреза гиалинового хряща из ребра кошки, 1 — основное вещество; 2 — клетки, помещающиеся в хрящевых полостях; 3 — хрящевые капсулы. Увелич. в 200 раз. Фиг. 9. Часть разреза сетчатого хряща из уха теленка. 1 — эластические волокна; 2 — хрящевые клетки, лежащие в полостях основного вещества (3) . Увелич. в 535 раз. Фиг. 10. Часть разреза волокнистого хряща из межпозвоночной связки теленка. 1 — пучки фибриллей; 2 — хрящевые клетки, помещающиеся в полостях основного вещества. Увелич. в 535 раз. Фиг. 11. Часть поперечного разреза из декальцинированной кости собаки. 1 — основное вещество, расположенное концентрическими пластинками вокруг Гаверсовых каналов (2); 3 — костные щели с лежащими в них костными клетками. Увелич. в 200 раз.
Волокнистая соединительная Т. (табл. II, фиг. 3 и 4) характеризуется тем, что основное вещество ее построено из различной толщины пучков (волокон) соединительнотканных фибриллей; последние связываются в пучки при помощи большего или меньшего количества бесструктурного и полужидкого склеивающего вещества. К пучкам прилегают звездчатой формы клетки собственно соединительной Т. и различного рода другие клеточные элементы. Кроме того, между пучками фибриллей всегда находятся еще особенные волокна — эластические волокна, являющиеся постоянными спутниками всякой волокнистой соединительной Т. Пучки фибриллей имеют различную толщину и представляются в виде более или менее сдавленных цилиндров или в форме лент. Каждый пучок состоит из тончайших соединительнотканных фибриллей, склеенных, как и в ретикулярной Т., незначительным количеством склеивающего вещества, которое легко растворяется в известных реактивах, напр., в пикриновой кислоте, и тогда пучки легко распадаются на отдельные фибрилли. От действия кислот и щелочей пучки быстро набухают и растворяются, а при варения они дают клей (глутин), почему их называют иначе клейдающими пучками. Кроме пучков соединительнотканных фибриллей, в состав всякой волокнистой соединительной Т. входят еще особенные волока, отличающиеся от соединительнотканных пучков сильною светопреломляющею способностью и свойством противостоять действию кислот и щелочей. Волокна эти носят название эластических и состоят из эластина. Означенные волокна дихотомически делятся и, соединяясь между собою, образуют сети с узкими или широкими петлями. Большинство клеток волокнистой соединительной Т. имеет форму тонких пластинок, от которых отходит много пластинчатых и ветвящихся отростков. В каждой клетке помещается одно круглое или овальной формы ядро, причем обыкновенно все клетки тесно прилегают к поверхности пучков фибриллей и составляют род клеточных футляров. Иногда клетки располагаются на поверхности пучков рядом одна возле другой и, соединяясь межклеточным веществом, образуют, как, напр., в сухожилиях, целые клеточные ленты. В тех случаях, когда пучки фибриллей тесно прилегают друг к другу и идут в одном каком-либо определенном направлении, на поверхности клеток оттискиваются от давления, оказываемого на них пучками, гребешки, присутствие которых придает клеткам своеобразную форму. Вообще можно сказать, что форма описываемых клеток зависит до известной степени от расположения пучков фибриллей волокнистой соединительной Т. Помимо указанных клеток, в волокнистой соединительной Т. находятся еще лейкоциты ("странствующие" или "блуждающие клетки") и особенные зернистые клетки, происхождение которых пока не вполне выяснено. Различают три формы клеток, характеризующихся присутствием в них зернышек. Одни из них имеют круглую или веретенообразную форму и состоят из слегка зернистой протоплазмы, в которой помещается большое круглое или овальное ядро, — это "плазматические клетки" Вальдейера. Другого рода клетки круглы или овальны; в протоплазме их много блестящих зернышек, способных окрашиваться анилиновыми красками; в протоплазме их располагаются круглые или овальные ядра, присутствие которых иногда маскируется означенными зернышками. Они названы Эрлихом "тучными клетками" и, подобно первым, чаще всего встречаются поблизости кровеносных сосудов. Третья форма клеток — "клазматоциты" Ранвье: большие клетки с перетянутыми местами отростками, от которых на местах перетяжек легко могут отделяться небольшие частички. Означенные клетки, по мнению Ранвье, при воспалении дают материал для образования гноя. По всей вероятности, все перечисленные формы зернистых клеток представляют собою лишь видоизменения белых кровяных клеток, вышедших из кровеносных сосудов в окружающую их волокнистую соединительную Т. Смотря по тому, в каком отношении друг к другу стоят пучки фибриллей, волокнистая соединительная Т. является то в виде рыхлой, или так наз. "бесформенной", то в виде плотной, или "форменной", соединительной Т. В рыхлой Т. пучки фибриллей различным образом пересекаются друг с другом (фиг. 3), вследствие чего между ними остаются более или менее значительные промежутки, заполненные бесструктурным, полужидким основным веществом. Этот вид волокнистой соединительной Т. входит в состав различного рода органов (печени, легких, стенки кишечного канала); из нее построен слой кожи, известный под названием подкожной жирной клетчатки, а равно Т., связывающая различные внутренние органы. В плотной соединительной Т. пучки фибриллей довольно тесно прилегают друг к другу и идут в одном каком-либо или в разных направлениях (фиг. 4) и составляют нередко целые органы, как, напр., сухожилия, фасции, надкостницу, многие связки и пр. В волокнистой соединительной Т. постоянно встречаются эластические волокна, а в клетках ее часто находятся различные включения в виде капелек жира и зернышек пигмента. Если только что указанные волокна или клетки с включениями находятся в волокнистой соединительной Т. в преобладающем количестве, то последняя меняет несколько свой характер, смотря по тому, преобладают ли в ней эластические волокна, клетки с жировыми каплями или с зернышками пигмента. В первом случае волокнистую Т. называют эластическою, во втором — жировой Т. и, наконец, в третьем — пигментной Т. d) Эластическая Т. (фиг. 5) состоит из эластических волокон, огромное количество которых обыкновенно примешивается к волокнистой соединительной Т. некоторых связок, а равно к Т. стенок артерий и вен, где волокна нередко сливаются вместе и образуют продырявленные пластинки, известные под названием эластических, или окончатых, пластинок. Волокна идут или параллельно, или же перекрещиваются между собою в разных направлениях, причем на своем пути они ветвятся и, соединяясь друг с другом, составляют эластические сети. Они имеют лентовидную или цилиндрическую форму и часто достигают толщины 10 μ. и более. Органы или части их, в состав которых входит эластическая соединительная Т., отличаются своею упругостью и прочностью и нередко приобретают слегка желтоватую окраску. е) Жировая Т. (фиг. 6) отличается от рыхлой волокнистой соединительной Т. лишь тем, что в состав ее входит множество клеток, заключающих в своей протоплазме капельки жира, почему они и носят название "жировых клеток". Т. эта встречается преимущественно в определенных местах животного организма: в области шеи, в окружности почек, в той части соединительной Т. кожи, которая известна под именем "подкожной жирной клетчатки", в сальнике и пр. Некоторые исследователи (Тольд) рассматривают жировую Т. как совершенно особый вид соединительной Т. Другие исследователи (Флеминг), наоборот, полагают, что всякая клетка волокнистой соединительной Т. при подходящих условиях питания может превратиться в жировую клетку, вследствие чего считают жировую Т. лишь за видоизменение волокнистой соединительной Т. Жировые клетки имеют обыкновенно круглую или овальную форму и в большинстве случаев состоят из одной или нескольких капелек жира, окруженных более или менее тонким слоем протоплазмы, в одном каком-либо месте которой располагается круглое, несколько сплюснутое ядро. Самый периферический слой протоплазмы имеет более плотную консистенцию и образует клеточную оболочку, присутствие которой препятствует слитию жировых капель соседних клеток. Если рассматривать жировые клетки в оптическом разрезе и при таком положении их, когда ядра находятся сбоку, то тогда слой протоплазмы, окружающий жировые капли, будет казаться в виде ободка, утолщенного в том месте, где лежит ядро. У зародышей и молодых животных обыкновенно в жировой клетке помещается несколько различной величины капелек жира, которые затем сливаются в одну большую каплю, оттесняющую протоплазму и ядро к периферии. Некоторые реактивы, как, напр., осмиевая кислота, окрашивают жир (в черный цвет) и дают возможность легко отыскать его в клетках даже в том случае, когда он находится в них в виде мелких капелек. При жизни жир имеет слегка желтоватую или оранжевую окраску. Вскоре после смерти жир, заключенный в клетках, изменяется и превращается в игольчатые кристаллы жирных кислот (пальмитиновой и стеариновой). У старых субъектов и вообще при недостаточном питании жир постепенно исчезает из клеток, вследствие чего последние уменьшаются в объеме — это так называемая "простая жировая атрофия". Нередко на место исчезнувших капелек жира появляется богатая слизью жидкость — такой процесс известен под названием "серозной жировой атрофии". Жировая Т. имеет значение регулятора в процессах питания и обмена веществ в организме; образуя довольно толстый слой внешних покровов тела (так назыв. подкожную жирную клетчатку), жировая Т. благодаря своей плохой теплопроводности защищает организм от больших потерь тепла и в то же время предохраняет его от различных внешних механических насилий. Заполняя промежутки между мышцами и пр., жировая Т. придает известным частям тела округленность форм. f) Пигментная ткань (фиг. 7). Если в волокнистой соединительной Т. много клеток с зернышками особенного красящего вещества, то она называется пигментной. Такие клетки находятся в большом количестве в соединительной Т. кожи сильно окрашенных рас, напр. негров, и у различного рода низших позвоночных (рыб, амфибий, рептилий), где они располагаются почти непосредственно под эпителиальным покровом кожи. Пигментные клетки обладают способностью выпускать и втягивать свои отростки, что непосредственно зависит от нервной системы. Новейшие наблюдения показали, что к означенным клеткам подходят нервные волокна, оканчивающаяся на поверхности клеток разветвлениями в виде густой сети.
Между исследователями все еще ведется спор, возникают ли пучки соединительнотканных фибриллей, а равно и эластические волокна из клеток эмбриональной Т. или же из бесструктурного сначала основного ее вещества. На основании новейших наблюдений Шульце, Флеминга, Спулера и др. вероятнее всего, что соединительнотканные фибрилли происходят из протоплазмы клеток эмбриональной Т. путем постепенной дифференцировки и расщепления ее на тончайшие ниточки. Что касается эластических волокон, то они также появляются сначала внутри клеток в виде мелких блестящих зернышек, которые затем сливаются в нити. Жировая Т., по мнению одних исследователей, образуется из клеток собственно-волокнистой соединительной Т., по мнению же других — возникает из особенных круглых, слегка зернистых клеток, похожих на плазматические клетки. Последние в том месте, где должна образоваться жировая Т., собираются в группы — так назыв. "первичные органы жировых долек". Затем в каждой клетке начинают постепенно появляться сначала мелкие капельки жира; последние все более и более увеличиваются в объеме и в конце концов сливаются в одну или в несколько больших капель, вследствие чего протоплазма с ядром оттесняются к периферии клетки и таким образом получается типичная жировая клетка. Собственно соединительная Т., в особенности рыхлая, и жировая Т. довольно богата кровеносными и лимфатическими сосудами. Последние начинаются в ней особенными, имеющими неправильную, часто звездчатую форму щелями (межтканевыми соконосными промежутками), связанными между собою при помощи тончайших канальцев, известных под названием "соконосных канальцев". Нервы оканчиваются в собственно соединительной Т. особенными чувствительными аппаратами, которые представляются в виде различной величины и формы древовидных разветвлений, помещающихся между пучками соединительнотканных фибриллей.
2) Хрящевая Т. отличается от собственно соединительной Т. гораздо большею плотностью и упругостью и при варении дает так назыв. хрящевой клей (хондрин), в состав которого входит глутин (см. выше) с примесью слизи (муцина). Она составляет переходную форму к костной Т. Хрящевая Т. или образует целые органы, или же является в виде различной величины и формы пластинок и, за немногими лишь исключениями, окружается довольно плотной соединительнотканною оболочкой, называемой "надхрящницей" (perichondrium). Обыкновенно хрящевая Т. встречается в форме гиалинового, сетчатого или упругого (эластического) и волокнистого хрящей. Все три указанные формы хрящевой Т. по своему строению очень немногим отличаются друг от друга. Гиалиновый хрящ (фиг. 8) покрывает суставные концы костей, из него построены хрящевые концы ребер, большинство хрящей гортани, хрящевые кольца, входящие в состав дыхательного горла и бронхов; наконец, большинство костей скелета в известный период жизни человека и млекопитающих состоит из гиалинового хряща. Он имеет молочно-белый цвет и кажется однородным, но под микроскопом нетрудно заметить, что он составлен из основного вещества с расположенными в нем клетками — "хрящевыми клетками". Основное вещество представляется однородным и бесструктурным, но если его обработать известными реактивами или исследовать гиалиновый хрящ у старых субъектов, то в таком случае ясно выступает состав основного вещества из тончайших соединительнотканных фибриллей, которые склеиваются ничтожным количеством так назыв. "межфибриллярного вещества". Так как в живом хряще фибрилли и склеивающее их вещество имеют один и тот же показатель преломления, то поэтому основное вещество кажется однородным. Обыкновенно все основное вещество изрыто круглой и овальной формы полостями — хрящевыми полостями, в которых помещаются такой же формы хрящевые клетки. Хрящевые полости, а следовательно, и клетки располагаются на известном расстоянии друг от друга по всему основному веществу, или же они лежат группами — по три, по четыре и больше вместе. Лишь у самой периферии хрящевых образований, вблизи надхрящницы, полости и клетки кажутся более или менее сдавленными и помещаются одна возле другой. Часть основного вещества, непосредственно окружающая и ограничивающая хрящевые полости, сильнее преломляет лучи света и гораздо интенсивнее окрашивается известными (напр. анилиновыми) красками, чем остальная часть основного вещества; она, кроме того, не разрушается от действия минеральных кислот (напр. соляной и серной кислот), а равно при мацерации хряща, и образует так назыв. хрящевые капсулы. В капсулах нередко можно видеть слоистость, т. е. они кажутся как бы составленными из нескольких следующих друг за другом слоев. Часто в одной капсуле или, вернее сказать, в ограничиваемой ею хрящевой полости располагаются 2, иногда 3—4 хрящевых клетки. Происхождение капсул объясняется историей развития хряща: они рассматриваются как продукт выделения клеток, который, становясь постепенно плотным и дифференцируясь, образует вокруг клеток род оболочки. Обыкновенно в эмбриональном хряще клетки помещаются близко одна от другой и отделяются друг от друга лишь тонкими оболочками — капсулами. Но вслед за образованием и уплотнением первой капсулы, каждою клеткою выделяется новое количество уплотняющегося по ее периферии вещества, образуется новая капсула, которая оттесняет раньше возникшую капсулу дальше к периферии. Затем точно таким же способом появляются все новые и новые капсулы, вследствие чего хрящевые клетки все более и более отодвигаются друг от друга, а капсулы получают слоистое строение. Самые наружные (периферические), а вместе с тем и самые старые по своему происхождению капсулы, сливаясь между собой, образуют основное вещество хряща, между тем как более молодые из них окружают еще отдельные клетки или группы последних. Присутствие нескольких клеток в одной полости происходит благодаря делению заключающейся в ней первоначально одной материнской клетки на две дочерних клетки, а затем благодаря последовательному делению этих последних, при чем нередко дочерние клетки отделяются друг от друга перегородками из основного вещества. Таким образом, каждая клеточная группа представляет собою одно клеточное семейство. Что касается хрящевых клеток, то они имеют круглую или овальную форму; только у немногих животных (некоторых рыб, головоногих и др.) хрящевые клетки принимают звездчатую форму. Обыкновенно каждая хрящевая клетка тесно прилегает к стенке хрящевой полости, отделяясь от нее лишь весьма узким вокругклеточным пространством, наполненным лимфою. Но при известных условиях, напр., от действия различного рода реактивов (алкоголя, хромовой кислоты и др.), клетки съеживаются и отделяются или целиком от стенок полостей и в таком случае получают вид небольших комочков, окруженных более или менее широким и светлым ободком, или же съеживаются неравномерно и местами прилегают к стенкам полостей; в последнем случае они являются в виде звездчатой формы образований. Хрящевые клетки лишены оболочки, причем протоплазма их имеет нитчатое строение, которое можно наблюдать даже на живых клетках. Нередко в протоплазме клеток встречаются различные включения, как, напр., капельки жира (у стариков и тучных субъектов), капли гликогена и пр. В каждой клетке помещается одно, иногда два ядра круглой или овальной формы; деление ядер происходит путем митоза. В старческом возрасте основное вещество хряща часто подвергается известным изменениям: оно обызвествляется, окостеневает или подвергается так наз. асбестовому перерождению. Процесс обызвествления заключается в том, что в веществе хрящевых капсул начинают отлагаться мелкие зернышки углекислой извести, присутствие которых придает (при падающем свете) обызвествленным местам хряща особенный белый цвет. Такому процессу особенно часто подвергаются хрящи дыхательного горла, ребер и др. Что касается окостенения гиалинового хряща, то оно происходит точно таким же образом, как и энхондральное образование кости (см. Развитие костей), и преимущественно встречается в хрящах гортани и дыхательного горла, причем начинается нередко с двадцатилетнего возраста у мужчин и с двадцатидвухлетнего возраста у женщин. Наконец, асбестовое перерождение хряща обусловливается появлением в основном веществе волокон, которые не имеют ничего общего с фибриллями, входящими в состав основного вещества. Волокна появляются сначала поблизости хрящевых капсул, затем распространяются постепенно дальше, захватывая все новые и новые участки основного вещества. Измененные указанным образом места хряща приобретают асбестовый блеск, откуда и самый процесс получил название "асбестового перерождения". Сетчатый, или упругий, хрящ (фиг. 9) имеет гораздо меньшее распространение в животном организме, чем гиалиновый хрящ, и встречается в ушных раковинах, в надгортаннике, в Евстахиевых трубах и пр. Он отличается от гиалинового хряща лишь тем, что основное его вещество, построенное так же, как и в гиалиновом хряще, пронизывается эластическими волокнами. Последние вступают в хрящ из надхрящницы в виде различной толщины волокон и, постепенно делясь, образуют в основном веществе густейшую сеть. Присутствие в описываемом хряще эластических волокон придает ему значительную крепость и упругость и более или менее желтоватую окраску. Наконец, волокнистый хрящ (фиг. 10) принадлежит к менее всего распространенным в организме формам хрящевой Т. и находится в межпозвоночных связках (Hg. intervertebralia), в сесамовидных косточках, на местах прикрепления сухожилий к костям и проч.
По своему строению он несколько походит на плотную соединительную Т. (напр., на сухожилия). Основу его образуют пучки соединительнотканных фибриллей, идущие параллельно друг другу или пересекающиеся под углом, причем между пучками остаются небольшие промежутки, занятые основным веществом гиалинового хряща. В этом веществе находятся хрящевые полости с характерными клетками, которые чаще всего располагаются по 1 или по 2 — редко группами. Только по присутствию капсул и заложенных в них клеток часто удается отличить этот вид хряща от волокнистой (плотной) соединительной Т.
Питание хрящевой Т. Хрящ, подобно всякой другой Т., нуждается для сохранения своей жизни в притоке к нему питательного материала. Ввиду того, что хрящ обыкновенно не содержит кровеносных сосудов, за исключением лишь тех мест, где происходит его рост или процесс окостенения, питание хрящевой Т. должно происходить каким-либо другим путем. Исследователи (Спина, Будге, Вольтерс и др.) уже давно пытались найти такие пути и пришли к тому заключению, что хрящевые полости (перицеллюлярные пространства) соединяются между собою целою системою тончайших соконосных канальцев, которые стоят в непосредственном сообщении с лимфатическими сосудами надхрящницы и доставляют хрящу необходимый питательный материал. Другие исследователи, наоборот, сомневаются в существовании подобного рода канальцев, хотя нужно заметить, что у некоторых животных, напр. у хрящевых рыб, такие канальцы имеются в действительности и могут быть обнаружены при помощи тех же способов, какие употребляются для их обнаружения в волокнистой соединительной Т. В каком отношении находятся нервы к хрящевой Т. — мы пока еще ничего не знаем.
Развитие хрящевой Т. происходит из клеток среднего зародышевого листка (мезодермы), причем зачатки этой Т. мы находим у человека на 6—7 неделе зародышевой жизни. Хрящевые клетки происходят из индифферентных сначала клеток мезодермы, а основное вещество хряща является уже продуктом, постепенно выделяемым молодыми хрящевыми клетками.
3) Костная Т. (фиг. 11) принадлежит к самым плотным и твердым Т. животного организма и служит опорой и защитой для более мягких его частей. В состав ее, как и в состав всякой соединительной Т., входит основное вещество и клетки, причем первое из них является пропитанным солями углекислой и фосфорнокислой извести, присутствие которых придает костной Т. известную твердость и устойчивость. Известковые соли легко могут быть удалены из костной Т. действием на нее неорганических или органических кислот (напр. соляной кислоты, хромовой или пикриновой кислот), после чего костная Т. становится мягкою и без труда режется ножом. Основное вещество, лишенное солей извести, по своему химическому составу вполне подходит к волокнистой соединительной Т. и состоит из так наз. оссеина, или костного хряща. Из костной Т. построены кости, наружную поверхность которых, за исключением их суставных концов, покрывает довольно плотная оболочка — надкостница (periosteum). В волокнистом основном веществе костной Т. имеется множество угловатой формы полостей, которые сообщаются между собою помощью тончайших ветвящихся канальцев — костных канальцев. В указанных полостях помещаются звездчатой формы клетки — костные клетки, образования, аналогичные клеткам других видов соединительной Т. Кроме того, все основное вещество костной Т. пронизывает целая сеть особенных каналов, известных под названием Гаверсовых каналов. Они с одной стороны открываются на периферии костей (каналы Фолькмана), с другой — в костномозговой канал (в трубчатых костях) и в полости губчатого вещества костей (в коротких и плоских костях); в них помещаются кровеносные сосуды и нервы костной Т. Подробности о строении и развитии костной Т. — см. Кость и Костная Т.
III. Мышечная ткань (табл. III). В состав этой Т. входят гладкие и поперечно-полосатые мышцы. Первые называются непроизвольными мышцами, так как сокращение их не подлежит нашей воле. Вторые называются произвольными мышцами, ибо сокращение их элементов подчиняется нашей воле. Между указанными двумя формами мышечной Т. существует еще переходная форма, к которой относятся мышцы сердца. Они по своему строению подходят ближе к поперечно-полосатым мышцам, а по функции, наоборот, должны быть причислены к гладким мышцам.
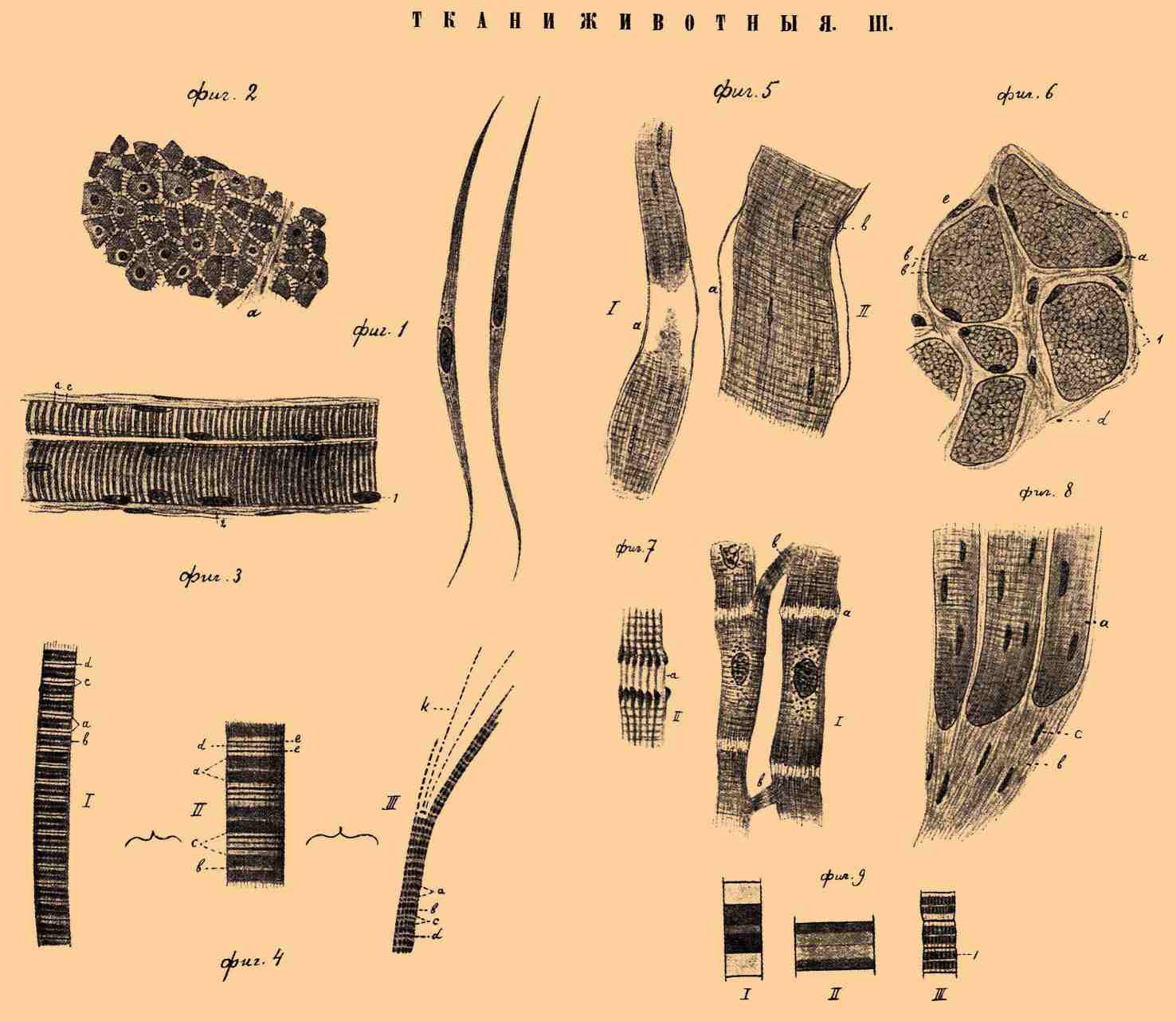
ТКАНИ ЖИВОТНЫЕ III.
Фиг. 1. Две изолированные клетки гладких мышц из кишечника кошки. Увелич. в 500 раз. Фиг. 2. Часть поперечного разреза кольцевидного слоя гладких мышц тонкой кишки собаки. Видны промежутки между поперек перерезанными мышечными волокнами и протоплазматические мостики, a — часть соединительнотканной прослойки. Увелич. в 500 раз. Фиг. 3. Части двух поперечно-полосатых мышечных волокон из языка человека. 1 — мышечные ядра; 2 — соединительнотканные прослойки между мышечными волокнами; a — толстый поперечный диск; с — светлый поперечный диск. Увелич. в 500 раз. Фиг. 4. I — часть поперечно-полосатого мышечного волокна мокрицы; II — часть мышечного волокна домашней мухи; III — частица расщипанного поперечно-полосатого мышечного волокна саламандры, а — толстый темный поперечный диск; b — срединный (Гензеновский) диск; с — тонкий светлый поперечный диск; d — межуточный (Меркелевский) диск; е — добавочные диски; k — мышечные фабрилли. Увелич. в 500 раз. Фиг. 5. Кусочки двух изолированных поперечно-полосатых мышечных волокон лягушки. а — сарколемма; b — мышечные ядра. Увелич. в 200 раз. Фиг. 6. Часть поперечного разреза портняжной мышцы собаки. 1 — поперечные разрезы мышечных волокон; а — мышечные ядра; b — поля Конгейма; b' — саркоплазма; с — сарколемма; d — соединительнотканные прослойки между мышечными волокнами; е — ядра соединительнотканных клеток. Увелич. в 500 раз. Фиг. 7. I — части двух мышечных волокон сердца человека. а — промежутки между мышечными клетками; b — отростки, отходящие от мышечных клеток. Увелич. в 500 раз. II — часть промежутка между 2-мя мышечными клетками; a — связующие ниточки, отходящие от утолщенных концов мышечных фибриллей. Сильное увеличение. Фиг. 8. Часть разреза на месте перехода поперечно-полосатой мышцы собаки в сухожилие. а — мышечные волокна; b — сухожилие; с — ядра клеток сухожилия. Увел. в 200 раз. Фиг. 9. Отдельные мышечные сегменты в расслабленном (I) и сокращенном (II) состоянии. III — три сегмента из поперечно-полосатого мышечного волокна осы (по Шефферу); 1 — канальцы, пронизывающие анизотропное вещество. Схематический рисунок.
а) Гладкие (непроизвольные) мышцы (фиг. 1 и 2) входят преимущественно в состав различного рода внутренних органов: желудочно-кишечного канала, мочевого пузыря, артериальных и венозных сосудов, многих желез и пр. Ткань гладких мышц состоит из длинных, веретенообразной формы и довольно толстых клеток, так назыв. гладких мышечных волокон. В состав тела каждой мышечной клетки входят тончайшие двоякопреломляющие свет ниточки — мышечные фибрилли (фиг. 1); они обыкновенно тянутся от одного до другого конца клетки и отделяются друг от друга незначительным количеством недифференцированной протоплазмы, или так называемый саркоплазмы (фиг. 1). В последней иногда находятся включения в виде зернышек пигмента и даже капелек жира. Особенной оболочки мышечные клетки не имеют. В средней, самой толстой части тела клетки помещается палочкообразное или овальное ядро с одним или несколькими ядрышками, причем саркоплазма скопляется около ядра в несколько большем количестве, чем в остальной части клетки. Обыкновенно мышечные клетки располагаются или отдельно, напр. в мелких артериях, или соединяются в пучки, которые часто составляют мышечную сеть. Образуя пучки, пластинки и пр., мышечные клетки так тесно примыкают одна к другой, что между ними остаются лишь едва заметные промежутки, заполненные незначительным количеством соединительной ткани. От каждой мышечной клетки отходят тонкие ниточки, так назыв. протоплазматические мостики, которые в настоящее время принимаются за образования искусственные, возникающие от действия реагентов на клетки. В тех случаях, когда мышечными клетками образуются толстые пучки и слои, обыкновенно группы клеток отделяются друг от друга более или менее толстыми прослойками из рыхлой соединительной Т. Если через пучок или слой мышечных клеток провести разрез так, чтобы мышечные клетки были разрезаны поперек (фиг. 2), то легко заметить, что на поперечных разрезах они кажутся в виде многоугольников различной величины. При этом в некоторых многоугольниках видны перерезанные в поперечном направлении ядра, в других же ядер нельзя заметить. Промежутки между многоугольниками выступают в виде тонких светлых линий, через которые проходят иногда упомянутые выше протоплазматические мостики (фиг. 2). Только что описанный вид мышечных клеток в поперечном их разрезе объясняется тем, что одни из них являются перерезанными в самой толстой, ядросодержащей своей части, между тем как другие попали в разрез на месте более или менее близком к одному из концов клетки. Светлые линии между многоугольниками обозначают тончайшие прослойки соединительной ткани, связывающей мышечные клетки. В течение всей жизни организма происходит постоянное умирание мышечных клеток и замещение их новыми, молодыми элементами, которые возникают из старых путем митотического деления. В организме человека и млекопитающих имеются органы, в которых происходит периодическое увеличение количества и размеров мышечных клеток. Что касается эмбрионального развития гладких мышц, то они происходят преимущественно из мезодермы и лишь весьма немногие из них развиваются из эктодермы или энтодермы.
b) Поперечно-полосатые (произвольные) мышцы (фигуры 3, 4, 5 и 6) являются более дифференцированными, чем гладкие мышцы, и входят в состав всех мышц скелета. Элементы, образующие эти мышцы, имеют вид более или менее длинных волокон, которые кажутся исчерченными в поперечном направлении темными и светлыми полосками (фиг. 3), вследствие чего они и названы поперечно исчерченными — рубчатыми — мышечными волокнами. В них можно заметить еще и продольную исчерченность. Каждое мышечное волокно представляется в форме несколько сдавленного цилиндра, концы которого заострены или притуплены. Иногда они тянутся во всю длину данной мышцы и достигают 12 стм и более, иногда, наоборот, волокна прерываются в различных местах данной мышцы. В большинстве случаев в длинных мышцах волокна длиннее, чем в коротких. Толщина отдельных мышечных волокон у человека = 30,6 — 65,7 μ, но у различных мелких животных (мышей, крыс и пр.) волокна, вообще, толще, чем у крупных; у молодых животных толщина их меньше, чем у взрослых. Есть мышцы (языка, брюшные и др.), в которых многие мышечные волокна расщепляются на несколько часто вновь делящихся веточек с закругленными или притупленными концами. Каждое мышечное волокно по своему развитию и строению представляет не что иное, как колоссальную многоядерную клетку, которая окружается снаружи особенной тонкой оболочкою, известной под назв. сарколеммы. Кроме последней, в состав мышечных волокон входят: тонкие, исчерченные в поперечном направлении нити — мышечные (первичные) фибрилли, склеивающая их недифференцированная часть протоплазмы — саркоплазма — и, наконец, расположенные в этой последней — мышечные ядра. Сарколемма (фиг. 5, а) представляется в виде тонкой бесструктурной прозрачной оболочки, которая тесно прилегает к веществу мышечного волокна и окружает каждое волокно на всем его протяжении. Оболочку эту можно ясно видеть, если взять небольшой кусок свежей мышцы и расщипать иголками на предметном стекле в капле физиологического раствора поваренной соли. На приготовленных таким образом препаратах сарколемма местами отслаивается от вещества мышечных волокон в виде пузырей или же вещество некоторых волокон разрывается, между тем как сарколемма, будучи значительно эластичнее мышечных фибриллей, остается в целости и обнаруживается на известном протяжении на местах разрывов. Означенной оболочки не имеют лишь мышечные волокна некоторых низших животных. Мышечные (первичные) фибрилли (фиг. 4, III к) имеют вид тонких нитей, вещество которых дифференцировано таким образом, что каждая фибрилля кажется как бы составленною из целого ряда правильно чередующихся и различно преломляющих свет частичек. Последние имеют приблизительно одинаковую толщину, причем одни из них сильно преломляют свет и при известной постановке трубы микроскопа представляются темными (фиг. 3 и 4, a), другие, наоборот, слабее преломляют лучи света и при той же постановке трубы являются светлыми (фиг. 3 и 4, с). Если рассматривать мышечные фибрилли в поляризованном свете, то окажется, что темные частицы являются двоякопреломляющими — анизотропными, а светлые — однопреломляющими свет, изотропными. В большинстве случаев различными красящими веществами анизотропные частицы окрашиваются интенсивнее изотропных. В состав каждого волокна входит множество мышечных фибриллей, а так как в каждой фибрилле темные и светлые частицы лежат на одном уровне с такими же частицами других соседних фибриллей, то все мышечное волокно должно казаться исчерченным в поперечном направлении темными и светлыми полосками, которые проникают через всю толщу волокна. Таким образом, кажущиеся полоски на самом деле производят впечатление дисков, расположенных друг над другом наподобие монет, сложенных в столбики. Рассматривая мышечные фибрилли при помощи сильных увеличений, нетрудно заметить, что каждая темная частица разделяется в средней своей части тонким слоем светлого однопреломляющего свет вещества (фиг. 4, b), а каждая светлая частица в свою очередь разделяется еще таким же тонким слоем темного, двоякопреломляющего свет вещества (фиг. 4, d). Означенные слои правильно чередуются и лежат на одном и том же уровне во всех мышечных фибриллях, составляющих данное мышечное волокно. Вследствие этого в мышечном волокне через середину темных полосок анизотропного вещества обыкновенно проходят узкие светлые полоски, а светлые полоски изотропного вещества разделяются узкими темными полосками анизотропного вещества; полоски эти, подобно вышеописанным, проникают через всю толщу мышечного волокна, т. е. представляются в виде дисков. Таким образом мышечные волокна кажутся как бы составленными из правильно чередующихся дисков: а) толстого анизотропного (фиг. 4, a), в середине которого находится b) тонкий изотропный диск (фиг. 4, b), и из с) толстого светлого — изотропного (фиг. 4, с), с проходящим через его середину d) тонким анизотропным диском (фиг. 4, d). Каждому из этих дисков дают особые названия: диск а называют толстым темным поперечными диском, b — срединным, или Гензеновским, диском, с — тонким светлым поперечным диском и d — межуточным, или Меркелевским, диском. Но, кроме перечисленных частиц, на которые дифференцируются мышечные фибрилли, а следовательно, и волокна, в мышцах многих низших животных (насекомых) можно констатировать, что в каждой половинке светлой, изотропной частицы помещается еще темный слой однопреломляющего свет вещества; слой этот на целом мышечном волокне кажется в виде двух темных, так называемых добавочных, дисков (фиг. 4, II е), расположенных в светлом толстом диске по обе стороны промежуточного диска. Все описанные частицы составляют одно неразрывное целое — одну мышечную фибриллу, дифференцировка которой на участки, обладающие различными физическими свойствами, есть лишь результат их функциональной деятельности. Под влиянием 93% алкоголя мышечные волокна, как это было впервые указано Боумэном, распадаются в поперечном направлении на диски, которые получили название Боумэновских дисков. От действия крепких кислот (напр. соляной кислоты) мышечные волокна также распадаются на диски, в состав которых входят промежуточный диск и, вероятно, части поперечных темного и светлого дисков. Во время сокращения мышечные волокна становятся короче и толще, а вместе с этим уменьшается толщина дисков и увеличивается ширина их. Обыкновенно анизотропное вещество становится более светлым, а изотропное, наоборот, преломляет лучи света сильнее. Это явление указывает, что во время сокращения часть изотропного вещества как бы переходит в анизотропное вещество. По наблюдениям Шёффера, сделанным над живыми мышцами осы, все анизотропное вещество (толстый поперечный диск) пронизывается тончайшими канальцами, которые простираются вплоть до срединного (Гензеновского) диска (фиг. 9). В эти канальцы при сокращении мышечных волокон вдавливается жидкое изотропное вещество и растягивает их, вследствие чего толщина диска уменьшается, а ширина увеличивается. Что касается саркоплазмы (фиг. 6, b'), то она представляется в виде однородного бесструктурного вещества, которое располагается более или менее тонким слоем под сарколеммой и оттуда проникает в толщу мышечного волокна, образуя в нем целый ряд тонких перегородок. Последние, таким образом, разбивают мышечные фибрилли на отдельные пучки, границы которых ясно выступают на поперечных разрезах мышечных волокон в виде тонких светлых линий. Означенные линии разделяют поперечный разрез мышечного волокна на мелкие, многоугольной формы участки (фиг. 6, b), внутри которых можно заметить присутствие более или менее мелких зернышек. Замечаемые на поперечных разрезах волокон участки суть не что иное, как поперечные разрезы пучков фибриллей, называемые полями Конгейма (фиг. 6, b), а зернышки в них — поперечные разрезы самых фибриллей. В мышечных волокнах членистоногих количество саркоплазмы бывает гораздо больше, чем в мышечных волокнах человека и млекопитающих, причем она образует довольно толстые прослойки. Последние разделяют волокно на несколько более крупных участков — пучков, которые в свою очередь тонкими прослойками саркоплазмы разбиваются на мелкие пучки фибриллей. Окружая пучки фибриллей, саркоплазма проникает в то же время и в промежутки между отдельными фибриллями, но лишь в весьма ничтожном количестве. Таким образом, количество саркоплазмы в мышечных волокнах низших животных бывает значительно меньше, чем в волокнах человека и млекопитающих. Под влиянием воды, разведенных кислот и щелочей саркоплазма растворяется. Обыкновенно в саркоплазме помещаются включения в виде мелких зернышек. Иногда, напр. в мышцах голодающих животных или при отравлении мышьяком, фосфором и пр., в саркоплазме появляются мелкие капельки жира, иначе говоря, происходит жировая дегенерация мышц. Саркоплазма, вероятно, играет важную роль в питании и размножении мышечных волокон, в ней также помещаются и мышечные ядра (фиг. 3 и 5 b; фиг. 6, a). Означенные ядра имеют овальную форму и расположены так, что продольная ось их совпадает с осью мышечного волокна. Обыкновенно ядра залегают в том месте мышечного волокна, где имеется большее скопление саркоплазмы, почему у человека и млекопитающих они в большинстве случаев лежат в периферическом слое саркоплазмы, непосредственно под сарколеммой. В мышечных волокнах низших позвоночных и насекомых ядра, наоборот, располагаются как у периферии, так и внутри волокна, в толстых прослойках саркоплазмы. Указанное расположение ядер особенно ясно выступает на поперечных разрезах мышечных волокон, окрашенных какими-либо ядерными красками — гематоксилином, борным и квасцовым кармином и др. У взрослых животных постоянно происходит, чисто физиологическим путем, умирание многих старых мышечных волокон и замещение их новыми, молодыми волокнами. Последние обыкновенно образуются из старых путем расщепления их в продольном направлении, причем в процессе образования новых волокон важную роль играют саркоплазма и мышечные ядра. Прежде всего, во внешнем слое саркоплазмы возникают путем митотического деления несколько рядов ядер (Вейсмановские волокна), а вместе с этим увеличивается также и количество саркоплазмы. Затем волокна разделяются в продольном направлении на известное количество новых волокон соответственно количеству рядов ядер. Что касается роста мышечных волокон в длину, то он, по-видимому, происходит на обоих концах каждого волокна и характеризуется увеличением здесь количества саркоплазмы и ядер.
Эмбриональное развитие поперечно-полосатых мышц у позвоночных происходит из мезодермы, причем клетки, дающие им начало, носят название "миобластов". Обыкновенно миобласты сначала представляются в виде одноядерных цилиндрической или веретенообразной формы клеток, протоплазма которых не имеет никакой определенной формы дифференцировки. Постепенно ядра этих клеток подвергаются митотическому делению и располагаются в осевой их части, а самые клетки удлиняются. Почти одновременно с этим периферический слой протоплазмы каждой клетки дифференцируется в мышечные фибрилли, между тем как центральная часть ее дифференцируется лишь впоследствии. Таким образом, одноядерная сначала клетка — миобласт — постепенно превращается в длинную клетку со многими ядрами, протоплазма ее дифференцируется в мышечные фибрилли, а остаток недифференцированной протоплазмы является в форме известной уже саркоплазмы. Сарколемма возникает лишь впоследствии и представляет собою образование, аналогичное клеточной оболочке.
с) Мышцы сердца (фиг. 7) занимают как бы середину между гладкими и поперечно-полосатыми мышцами, имея много общего как с теми, так и с другими. Несмотря на то что сокращение их не подлежит нашей воле, тем не менее, они имеют ясно выраженную поперечную исчерченность. Означенные мышцы состоят из одноядерных, веретенообразной (у низших позвоночных) и цилиндрической формы клеток, от которых отходят под разными углами короткие отростки (фиг. 7, b). Клетки эти располагаются рядами, причем концы одной клетки соединяются с концами соседних с нею клеток при помощи особенного склеивающего вещества (фиг. 7, а), вследствие чего возникают более или менее длинные волокна. Но, помимо того, боковые отростки клеток, составляющих данное волокно, точно таким же образом соединяются с отростками других соседних волокон. Благодаря существованию указанной взаимной связи между мышечными волокнами возникает мышечная сеть, или плетенка, составленная из множества склеенных друг с другом мышечных клеток. Ядра отдельных клеток помещаются в середине каждой клетки (фиг. 7), имеют круглую или овальную форму и обыкновенно окружаются небольшим количеством недифференцированной протоплазмы; вся остальная часть последней дифференцируется в мышечные фибрилли, разделенные лишь небольшим количеством саркоплазмы на пучки. Последние, как это легко видеть на поперечных разрезах волокон, в периферической части каждой клетки имеют в большинстве случаев лентовидную форму, а ближе к ее центру — многоугольную (полигональную) форму. Первые на поперечных разрезах являются в виде узких зернистых полосок, идущих радиально к центру клетки. Мышечные фибрилли имеют точно такое же строение, как и фибрилли поперечно-полосатых волокон. Обыкновенно концы фибриллей кажутся утолщенными и при помощи очень тонких ниточек соединяются с подобными же утолщениями фибриллей других соседних клеток. Означенные связующие ниточки (фиг. 7, II a) проникают через склеивающее вещество и до известной степени напоминают собою так назыв. протоплазматические мостики эпителиальных клеток. В отличие от произвольных мышечных волокон мышечные клетки сердца не окружены особенной оболочкой, т. е. не имеют сарколеммы. Подобно мышцам скелета, мышечные волокна сердца разделяются прослойками из рыхлой соединительной Т. на отдельные пучки различной величины и формы. Помимо того, от означенных прослоек отходят еще более тонкие перегородки в промежутки между мышечными волокнами и отделяют их друг от друга так же, как и в. поперечно-полосатых мышцах скелета.
Кровеносные сосуды. Мышечная Т., являясь одною из деятельнейших Т. животного организма, нуждается в постоянном притоке питательного материала и поэтому снабжена значительным количеством кровеносных сосудов; в этом легко можно убедиться, если наполнить сосуды какой-либо мышцы окрашенною массою, а затем приготовить микроскопические препараты. Обыкновенно уже макроскопически можно заметить, что к каждой мышце подходит несколько более или менее крупных артерий, которые постепенно делятся и вступают в толстые соединительнотканные прослойки мышц, где ими образуются широкопетлистые сети. От этих сетей отходят мелкие артерии в тонкие соединительнотканные перекладины и, разветвляясь в них, составляют новые сети, оплетающие тонкие пучки мышечных волокон. Означенные сети дают начало капиллярам, которые проникают в промежутки между отдельными мышечными волокнами и оплетают волокна в виде густейшей сети; петли последней представляются более или менее вытянутыми в длину. Из этой сети получают начало мелкие вены, которые сопровождают артерии и располагаются по одной с каждой стороны артериального сосуда.
Лимфатические сосуды мышц еще мало изучены. Они начинаются из лимфатических щелей и канальцев и направляются вместе с кровеносными сосудами.
Нервы. Мышечная Т. богата не только кровеносными и лимфатическими сосудами, но и нервами, которые оканчиваются в них особенными двигательными и чувствительными аппаратами.
Переход мышц в сухожилия (фиг. 8). Все мышцы прикрепляются к плотным или более мягким частям тела, как, напр., кости, Т. кожи и др., при помощи плотной соединительной Т., которая образует так назыв. сухожилия (см. подр. Поддерживающая соединительная Т.). Обыкновенно у обоих концов мышцы на местах прикрепления ее к сухожилиям мышечные волокна оканчиваются заостренными или притупленными концами, причем пучки мышечных фибриллей в каждом волокне оканчиваются не на одном уровне: одни из них оканчиваются раньше, другие позднее, что, конечно, зависит от формы концов самого мышечного волокна (фиг. 8). В том месте, где концы мышечных волокон упираются в сухожилия, в этих последних образуется целый ряд более или менее глубоких вдавлений. Пучки соединительнотканных фибриллей сухожилия, подойдя к мышце, рассыпаются на тонкие пучочки, которые вступают в промежутки между мышечными волокнами и переходят непосредственно в так назыв. perimysium internum (фиг. 6, d), т. е. в соединительную Т., отделяющую мышечные волокна друг от друга. Таким образом сухожилие вступает в тесную связь с соединительною Т., окружающею отдельные мышечные волокна данной мышцы. В сухожилиях помещаются кровеносные и лимфатические сосуды и нервы. Кровеносные сосуды располагаются сравнительно в незначительном количестве в прослойках из рыхлой соединительной Т., разбивающей сухожилия на пучки 2, 3 и т. д. порядков. Лимфатические сосуды, начинаясь соконосными щелями, помещающимся в промежутках между пучками 1-го порядка сухожилий и в прослойках из рыхлой соединительной Т., образуют на поверхности сухожилий густую сеть. Нервы оканчиваются в сухожилиях особенными нервными концевыми аппаратами в форме древовидных разветвлений и в виде небольших Фатер-Пачиниевых телец.
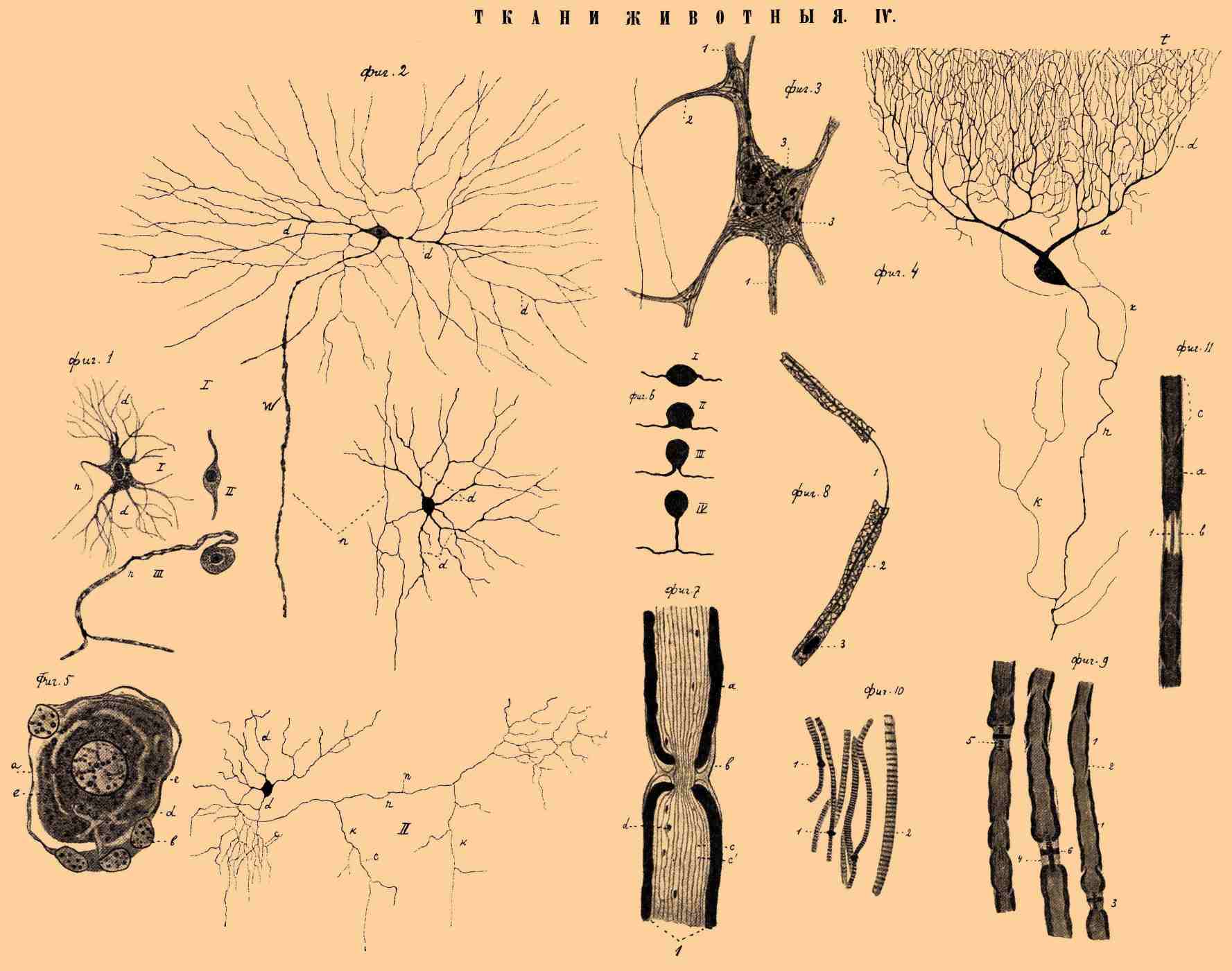
ТКАНИ ЖИВОТНЫЕ IV.
Фиг. 1. Нервные клетки: I — многоотростчатая клетка из спинного мозга теленка; II — двуотростчатая клетка из спинномозгового узла окуня; III — одноотростчатая клетка из спинномозгового узла кошки, n — нервный отросток; d — дендриты. Увелич. в 80 раз. Фиг. 2. Нервные клетки: I — типа Дейтерса (из сетчатки совы), II — типа Гольджи (из мозжечка стерляди) и III — нервная клетка третьего типа (из мозжечка собаки), n — нервный отросток; d — дендриты; k — коллатерали (боковые веточки). Увелич. в 80 раз. Фиг. 3. Многоотростчатая нервная клетка из сетчатки совы; виден состав тела клетки и ее отростков из фибриллей и межфибриллярного вещества, в котором помещаются тигроидные тельца (3). 1 — дендриты; 2 — нервный отросток. Увелич. в 440 раз. Фиг. 4. Клетка Пуркинье из мозжечка собаки. d — дендриты; n — нервный отросток; k — коллатерали; t — телодендрия. Увелич. в 240 раз. Фиг. 5. Спинномозговая клетка кролика. а — оболочка (капсула); b — ядра клеток, выстилающих внутреннюю поверхность оболочки; с — ядро; d — тигроидные тельца; е — внутриклеточные канальцы. Увелич. в 1000 раз (По Holmgren'y). Фиг. 6. Спинномозговая двуотростчатая клетка зародыша (I), постепенно превращающаяся во время дальнейшего развития организма (II, III) в одноотростчатую клетку. Схематический рисунок. Фиг. 7. Часть мякотного нервного волокна. а — мякотная — миелиновая оболочка; Шваннова оболочка на месте перехвата Ранвье; с — нервные фибрилли; с' — межфибриллярное вещество; d — тигроидные тельца. 1 — осевой цилиндр. Схематический рисунок. Фиг. 8. Мякотное нервное волокно лягушки, обработанное смесью спирта с эфиром и окрашенное гематоксилином. 1 — осевой цилиндр; 2 — неврокератиновая оболочка; 3 — ядро клетки Шванновой оболочки. Увелич. в 240 раз. Фиг. 9. Мякотные нервные волокна лягушки, обработанные осмиевой кислотой, 1 — сегменты и (2) насечки — вырезки — Лантерманна; перетяжка Ранвье; 4 — Шваннова оболочка; 5 — осевой цилиндр с поперечною исчерченностью и склеивающее вещество, которое вместе с осевым цилиндром дает фигуру креста. Увелич. в 440 раз. Фиг. 10. Осевые цилиндры нервных волокон из спинного мозга быка. 1 — утолщения на местах перетяжек и (2) Фроманновские линии. Увелич. в 440 раз. Фиг. 11. Мякотное нервное волокно лягушки. a — мякоть — миэлин; b — Шваннова оболочка; с — сегмент Лантерманна. 1 — осевой цилиндр. Увелич. в 500 раз.
IV. Нервная Т. (табл. IV и V) по той роли, какую она играет в жизни организма, можно сказать, занимает первое место среди всех остальных Т. В состав ее входят нервные клетки и нервные волокна; последние на самом деле являются лишь отростками нервных клеток и представляют вместе с ними одно неразрывное целое. В большинстве случаев от каждой нервной клетки отходит много отростков, из которых один непосредственно превращается в нервное волокно и получает название нервного (осево-цилиндрического) отростка (фиг. 1, 2, 3 и 4 n), остальные же — многократно делятся на множество веточек и составляют так назыв. дендриты (фиг. 1, 2, 3 и 4, d; протоплазматические отростки) клетки. Многие исследователи (Кахаль, Вальдейер, Ленгоссек и друг.), изучая отношение нервных клеток друг к другу, пришли к тому заключению, что они представляют собою совершенно самостоятельные, независимые друг от друга единицы, не связанные непосредственно с другими подобными же единицами — нервными клетками, — и дали им название "невронов". Отсюда возникло в последнее время учение "о невронах", которое вскоре приобрело себе много поклонников. Но в новейшее время благодаря работам А. Догеля, Апати, Бете и друг. исследователей учение о невронах теряет под собою почву и уступает место учению о тесной и непосредственной связи клеток между собою. Кроме нервных клеток со всеми их отростками, в состав нервной Т. входят еще элементы, образующие остов этой Т. и служащие для поддержки собственно нервных элементов. Остовом этим в центральной нервной системе является так наз. "невроглия" или "глия", а в периферической системе (в нервных стволиках, в симпатических и спинномозговых ганглиях) — фибриллярная соединительная Т.
Нервные клетки (фиг. 1, 2, 3 и 4) входят в состав центральной и периферической нервной системы и имеют многоугольную, круглую, грушевидную, овальную и веретенообразную форму. Величина клеток в разных отделах центральной нервной системы и у различных животных подвержена значительным колебаниям, и рядом с клетками, диаметр которых не превышает 0,004 мм, имеются клетки почти заметные невооруженным глазом — с диам. в 0,135—0,2 мм. К мелким клеткам относятся, напр., так наз. "маленькие клетки Рольджи" в зернистом слое мозжечка, многие клетки нервной оболочки глаза и др.; к большим клеткам принадлежат, между прочим, двигательные клетки передних рогов спинного мозга и вообще нервные клетки низших позвоночных, напр. рыб. Уже давно, еще в 1844 г., Ремак обратил внимание на то, что в состав тела нервных клеток входят тончайшие фибрилли. Наблюдения Ремака были подтверждены и значительно дополнены М. Шульце в 1871 г., а в последние 10 лет благодаря улучшению микроскопов получена возможность проникнуть глубже как в строение нервных клеток, так равно и нервных волокон. В настоящее время можно сказать, что в состав тела нервных клеток входят: a) хроматофильное, тигроидное вещество (фиг. 3, з), или так наз. тельца Ниссля; b) ахроматиновые нервные фибрилли (фиг. 3) и, наконец, с) ахроматиновое, аморфное основное вещество (фиг. 3). Ниссль, окрашивая нервные клетки различными, преимущественно основными, анилиновыми красками, заметил, что в протоплазме их располагаются небольшие, многоугольной формы образования, которые окрашиваются весьма интенсивно указанными выше красками и придают телу нервной клетки пятнистый вид или вид тигровой шкуры, откуда эти образования и получили название "тигроидных телец" (ф. 3, з). Они находятся не только в протоплазме клетки, но и в клеточных отростках — дендритах и нервном отростке. Каждое тигроидное тельце в свою очередь состоит из собрания мелких зернышек, связанных небольшим количеством неокрашивающегося вещества. Метиленовая синь и нейтральрот (А. Догель и Беккер) окрашивают зернышки тигроидных телец в живых нервных клетках, а это указывает, что они являются образованиями, постоянно присущими клеткам, и не возникают в них искусственно вследствие обработки клеток теми или другими реактивами. Тигроидные тельца в различных стадиях деятельности одной и той же клетки изменяются характерным образом; они подвергаются также изменению от действия на клетки разных ядовитых веществ, напр. свинца, мышьяка, стрихнина и др., а равно при отравлении организма продуктами обмена веществ известных микроорганизмов, напр. чумных бактерий и пр. Вещество, в котором помещаются тельца Ниссля, как это показали исследования Флеминга, А. Догеля, Рамон и Кахаля, Апати, Бете и др., состоит из множества тончайших ниточек — нервных фибриллей, — которые проходят в промежутках между тельцами, перекрещиваются в разных направлениях и, не анастомозируя друг с другом, вступают как в дендриты, так и в нервный отросток каждой клетки. Нервные фибрилли то идут пучками, то отдельно, то волнообразно изогнуты и являются самыми существенными элементами нервной клетки. Между нервными фибриллями и тигроидными тельцами располагается незначительное количество аморфного, однородного ахроматинового основного вещества. Кроме того, клетки спинномозговых и симпатических ганглиев у всех позвоночных окружаются еще тонкою, бесструктурною соединительнотканною оболочкою — капсулою, которая является лишь продолжением так наз. Шванновой оболочки нервных волокон (см. ниже). Оболочка эта в симпатических клетках млекопитающих продолжается и на толстые дендриты, а в спинномозговых клетках — на их единственный отросток. Внутренняя поверхность оболочки обыкновенно выстлана плоскими клетками эпителия, а между ним и телом клетки остается узкое щелевидное пространство, наполненное лимфою, почему его и наз. перицеллюлярным лимфатическим пространством (фиг. 5). В теле нервных клеток помещается в большинстве случаев круглое или овальной формы ядро (фиг. 1, 2, 3 и 5), которое лежит то в центре, то эксцентрически и заключает в себе одно или несколько ядрышек. Наконец, внутри тела многих нервных клеток, по наблюдениям Е. Гольмгрэна и др., располагается целая сеть тончайших канальцев, которые, по всей вероятности, сообщаются с перицеллюлярным лимфат. пространством и служат для проведения к клеткам питательных веществ и для удаления продуктов обмена веществ. Нередко в определенных нервных клетках имеются включения в виде зернышек бурого и желтого пигмента. От тела нервных клеток отходят много, или же только два и даже один, отростков. Все нервные клетки разделяют на три больших группы: на так называемые многоотростчатые, многополюсные (мультиполярные) нервные клетки (фиг. 1, I), двуотростчатые — биполярные (фиг. 1, II) (двухполюсные) и одноотростчатые — униполярные (фиг. 1, III) (однополюсные) клетки. К первой группе принадлежат большинство клеток центральной нервной системы (головного и спинного мозга), причем количество отростков может простираться от 3—4 до 6—10 и больше. К двуотростчатым клеткам относятся клетки спинномозговых ганглиев рыб и всех позвоночных в эмбриональном периоде их жизни, а равно клетки спирального узла (gangl. spirale) и так называемые обонятельные клетки. Наконец, одноотростчатые клетки входят в состав спинномозговых ганглиев человека и млекопитающих и, кроме того, имеются в симпатических ганглиях амфибий. Обыкновенно каждый отросток большинства спинномозговых клеток, пройдя известное расстояние, делится V- или Т-образно на две ветви (фиг. 1, III) из которых одна, более толстая ветвь идет в виде нервного волокна к периферии, а другая, тонкая ветвь направляется к центру (в спинной мозг) (фиг. 1, III). История развития этих клеток показывает, что в эмбриональной жизни они имеют биполярную форму, а затем постепенно, по мере роста клеточного тела, оба отростка приближаются друг к другу и наконец сливаются в один отросток, вследствие чего клетка из дву- становится одноотростчатой (фиг. 6). Кроме того, весьма вероятно, что существуют еще нервные клетки без отростков, или аполярные. К ним должны быть причислены особые клетки, входящие в состав чувствительных аппаратов, известных под названием телец Грандри, а также чувствительные клетки Меркеля. В нервных клетках в большинстве случаев один отросток превращается в нервное волокно и называется нервным отростком, остальные более или менее сильно ветвятся и носят название дендритов. Нервный отросток (осево-цилиндрический отросток, неврит; фиг. 1, 2, 4, n и фиг. 3, 2) начинается коническим утолщением от тела клетки или одного из ее дендритов, затем суживается и вскоре опять принимает вид более или менее толстого волокна, которое кажется гладким, довольно блестящим и нередко достигает длины более одного метра. В последнем случае нервные отростки переходят из одного отдела в другие отделы центральной нервной системы или же выходят за пределы этой системы, например нервные отростки двигательных клеток спинного мозга окружаются особенными оболочками и в таком виде получают название нервных волокон. Образования, которые мы обыкновенно называем нервами, нервными стволиками представляют собою не что иное, как собрание нервных отростков клеток. Пройдя более или менее значительное расстояние, смотря по типу нервной клетки, нервный отросток затем распадается на известное количество повторно делящихся концевых веточек. Во время своего хода нервный отросток отдает от себя то короткие, то, наоборот, длинные тонкие боковые веточки, так назыв. коллатерали (фиг. 2, III k и фиг. 4, k), которые оканчиваются таким же образом, как и нервный отросток.
Дендриты (протоплазматические отростки, фиг. 1, 2, 4, d и фиг. 3, 1) отходят от углов — полюсов — клеточного тела, имеют различную длину и толщину, слабее преломляют свет, чем первый отросток, и обыкновенно уже вблизи клетки начинают постепенно делиться под разными углами на большее или меньшее количество веточек. Последние вновь распадаются на еще более тонкие, вновь делящиеся концевые ниточки, или так назыв. телодендрии (telodendria) (фиг. 4, t). В большинстве случаев от нервной клетки отходит лишь один нервный отросток и много дендритов, напр. во многих клетках центральной нервной системы, или же один нервный отросток и один дендрит, как, например, в двуотростчатых клетках спинномозговых ганглиев рыб. В таких клетках, как одноотростчатые клетки спинномозговых узлов млекопитающих, отходящий от них отросток является смешанным: он V- или Т-образно (см. выше) делится на две ветви, причем та ветвь, которая идет к периферии, — представляет собою дендрит, а ветвь, направляющаяся к центру, — нервный отросток клетки (фиг. 1, III). Но имеются нервные клетки, от которых отходит не один, а несколько нервных отростков, напр. известные клетки сетчатки, или же клетки, не имеющие, по-видимому, вовсе нервных отростков; к последним принадлежат так назыв. диффузные амакриновые клетки сетчатки. Нервные отростки развиваются раньше дендритов, но и те и другие одинаково необходимы для жизни и деятельности каждой клетки. Принимая во внимание характер нервного отростка, принято различать несколько основных, типичных форм нервных клеток, между которыми, конечно, существуют переходные формы. К таким основным формам относят три типа: клетки с одним длинным отростком, который на известном расстоянии от клетки окружается оболочками и превращается в настоящее так назыв. мякотное или безмякотное нервное волокно, смотря по характеру оболочек. От него могут отходить боковые ветви (коллатерали), а затем он, пройдя обыкновенно большое расстояние, распадается на свои концевые веточки (фиг. 2, I). Клетки подобного рода относятся к типу так назыв. Дейтерсовых клеток, к которым принадлежат, напр., двигательные клетки спинного мозга, многие симпатические клетки и пр. Ко второму типу причисляют клетки с более или менее тонким нервным отростком, который проходит, сравнительно с клетками типа Дейтерса, небольшое расстояние и в большинстве случаев не выходит за пределы той части центральной нервной системы, в которой помещается данная клетка. Нервный отросток этих клеток отдает на всем своем протяжении боковые веточки, не окружается особенными оболочками и затем оканчивается многочисленными разветвлениями (фиг. 2, II). Как на пример подобных клеток, можно указать на так назыв. коробовые клетки мозжечка, на звездчатые клетки сетчатки и др. Наконец, третий тип клеток составляют клетки Гольджи, от которых отходит один тонкий и короткий нервный отросток; последний, не превращаясь в нервное волокно и не выходя за границы известного отдела нервной системы, распадается вблизи клеточного тела на множество постепенно делящихся тонких нервных ниточек (фиг. 2, III). К означенным клеткам относятся, напр., особенные клетки, располагающиеся в зернистом (ядерном) слое мозжечка и др. Нервная клетка со всеми ее отростками — дендритами и нервным отростком — представляет собою одно неразрывное целое, жизнь которого неминуемо нарушается, раз только будет нарушена целость одной из его составных частей. Поэтому нет никакого основания той или иной из них приписывать преобладающее значение. В пользу этого говорят наблюдения Валлера, сделанные еще в 1852 г., и новейшие исследования Ранвье, Фореля и в особенности Ван Геухтена, которые указывают, что нервное волокно подвергается дегенеративным процессам, как скоро будет нарушена связь его с нервною клеткою. Мало того, с перерезкою нервного волокна не только дегенерируется периферический его отрезок, но вместе с тем замечаются известные изменения и в теле той нервной клетки, от которой берет начало данное нервное волокно. Некоторые исследователи (Гольджи, Нансен) предполагали, что дендриты и тело нервной клетки играют чисто питательную роль. Далее, в последнее время Бете и друг. наблюдатели думали, что, напр., в явлениях рефлекса тело клетки не имеет никакого существенного значения. Но, тем не менее, не подлежит сомнению, что физиологические импульсы воспринимаются разветвлениями дендритов, передаются при посредстве их телу клетки, где они, вероятно, видоизменяются известным образом, а затем уже от тела клетки отводятся нервным ее отростком. Вопрос об отношении нервных клеток друг к другу до настоящего времени еще нельзя считать решенным окончательно. С точки зрения Рамон-Кахаля и его последователей, считающих каждую нервную клетку за отдельную анатомическую единицу — неврон, нервные клетки не соединяются непосредственно между собою, они только тесно соприкасаются друг с другом при помощи концевых разветвлений своих отростков. Но, тем не менее, уже сам Кахаль признает, что в отдельных, исключительных случаях существуют непосредственные анастомозы между отростками различных нервных клеток, напр. у многих беспозвоночных (насекомых), в симпатической нервной системе позвоночных и пр. Наблюдения А. Догеля, Апати, Бете и многих других исследователей показывают, что между разветвлениями клеточных отростков существует непосредственная связь, а не простое только соприкосновение. Указанные исследователи утверждают, что нервные фибрилли, самые существенные при передаче и воспринятии импульсов элементы, лишь проникают через тела нервных клеток, не начинаясь и не оканчиваясь в них. Фибрилли идут непрерывно от периферии к центру и от центра к периферии и представляют собою вполне замкнутую систему. Связь клеток одного вида с клетками другого вида происходит таким образом, что разветвления нервных отростков (фибрилли), оплетая дендриты и тела клеток, анастомозируют с фибриллями, входящими в состав последних. Во всяком случае, как это справедливо замечает Ферворн, на основании новейших исследований не подлежит сомнению, что у взрослых животных нервные клетки во многих случаях стоят в тесной и прямой связи друг с другом при помощи дифференцированного в фибрилли живого вещества их.
Нервные волокна. Нервные отростки многих клеток, как было указано выше, окружаются особенными оболочками и тогда получают название нервных волокон. Обыкновенно различают мякотные, или миелиновые, и безмякотные нервные волокна. Но, помимо того, встречаются волокна, лишенные оболочек — тогда их назыв. голыми осевыми цилиндрами. Мякотные (миелиновые или двуконтурные) волокна (фиг. 7, 8, 9 и 11) достигают от 0,001—0,02 мм в диаметре и состоят, как и всякое вообще нервное волокно, из осевого цилиндра, т. е. нервного отростка клетки, особенной мякотной — миелиновой — оболочки, из Шванновой оболочки, или неврилеммы и, наконец, из так назыв. эндоневральной, или Генлевской, оболочки. Если рассматривать мякотные волокна в свежем, живом виде, то они обыкновенно являются более или менее блестящими и резко очерченными с каждой стороны двуконтурною блестящей линией, откуда получили назв. двуконтурных волокон. В осевой части каждого такого волокна, как это особенно ясно видно на поперечном его разрезе, помещается то более, то менее толстая, иногда сплющенная нить — осевой цилиндр (фиг. 7, 8, 11, I). На свежих препаратах он кажется однородным и блестящим, на самом же деле, являясь нервным отростком клетки, имеет такое же строение, как и этот последний, т. е. состоит из тончайших ниточек — нервных, или первичных, фибриллей (фиг. 7, с) и весьма незначительного количества межфибриллярного вещества, или невроплазмы (фиг. 7, с'). Нервные фибрилли принадлежат к самым важным составным частям осевого цилиндра. Они идут, слегка изгибаясь волнообразно, и при нормальных условиях представляются совершенно гладкими. Межфибриллярное вещество — невроплазма — помещается в весьма незначительном количестве между фибриллями, а по всей периферии осевого цилиндра образует более или менее тонкий слой, составляя род оболочки вокруг осевого цилиндра. В межфибриллярном веществе находится незначительное количество мелких зернышек и глыбок (фиг. 7, d), вполне аналогичных тем включениям, которые под названием тигроидных телец имеются как в однородном основном веществе тела, так равно и в межфибриллярном веществе дендритов нервных клеток. Непосредственно за осевым цилиндром следует мякотная, или миелиновая, оболочка (фиг. 7 и 11, a; фиг. 9, 1), составленная из особенного жироподобного вещества — миелина. Вещество это кажется однородным, сильно преломляет свет, полужидко и окрашивается осмиевой кислотой, подобно жиру, в черный цвет. Оно растворяется в эфире, в смеси спирта с эфиром, крайне легко изменяется, обнаруживая так называемые явления свертывания, а именно миелин собирается в самой причудливой формы капельки. Толщина этой оболочки весьма различна: в толстых волокнах она окружает осевой цилиндр в виде толстого слоя, который на оптическом разрезе волокон является в форме двуконтурной линии, резко ограничивающей очертание каждого волокна; в тонких волокнах мякотная оболочка имеет столь незначительную толщину, что отчетливо видна лишь после обработки их осмиевой кислотой. Обыкновенно мякотная оболочка не окружает в виде непрерывного слоя все волокно от его начала до конца: она исчезает на более или менее близком расстоянии волокна от тела клетки и от места распадения его на концевые разветвления. Кроме того, и на всем том своем участке, который покрыт мякотью, последняя прерывается на определенных местах; означенные места известны под назв. перехватов или перетяжек Ранвье (фиг. 9, 3), отстоящих друг от друга на определенном расстоянии и разделяющих всю мякотную оболочку на отдельные сегменты. Длина этих сегментов постепенно уменьшается по направлению к концу нервного волокна, т. е., иначе говоря, перехваты Ранвье постепенно приближаются друг к другу. Таким образом, на месте каждого перехвата имеется лишь осевой цилиндр и так назыв. Шваннова оболочка, которая, однако, отделяется от осевого цилиндра то более, то менее толстым слоем особенного склеивающего вещества (фиг. 7). Последнее имеет вид кольца, через которое проходит осевой цилиндр. Если мякотные волокна обработать слабым раствором азотнокислого серебра и затем подвергнуть препарат действию солнечного света, то склеивающее вещество окрашивается в темно-бурый, почти черный цвет и кажется тогда в виде бурой или черной поперечной линии. Не трудно убедиться, что означенная линия на самом деле представляет кольцо, окружающее осевой цилиндр. Обыкновенно на местах перетяжек азотнокислым серебром окрашивается не только склеивающее вещество, но и самый осевой цилиндр, вследствие чего на указанных местах получаются фигуры, похожие на латин. крест; продольную часть последнего образует окрашенный серебром осевой цилиндр, а поперечную часть составляет склеивающее вещество. Фигуры эти были впервые описаны Ранвье и носят поэтому название крестов Ранвье (фиг. 9). Очень нередко осевой цилиндр окрашивается серебром на более или менее значительном протяжении от перехватов, причем в подобных случаях на нем замечается ряд поперечных бурых или черных линий. Означенные линии носят название Фромановских линий (фиг. 9, 5 и фиг. 10). Последние представляют собою образования искусственные, возникшие от действия азотнокислого серебра, и получаются на пучках соединительнотканных фибриллей, на дендритах нервных клеток и пр. В каждом сегменте нервного волокна, заключенном между двумя перехватами, миелиновая оболочка в свою очередь разделяется на небольшие участки, которые имеют вид воронок или конусов, известных под названием Шмидт-Лантермановских сегментов (фиг. 9, 1 и фиг. 11 с). Края последних представляются несколько утонченными и загнутыми внутрь или отогнутыми кнаружи, причем сегменты соединяются между собою таким образом, что суженный конец одного является немного вдвинутым в широкий конец другого сегмента и т. д. При этом образуются узкие кольцевидные щели, которые видны особенно ясно после обработки нервных волокон осмиевой кислотой, когда самые конические отрезки мякоти кажутся черными, а промежутки между ними имеют вид косых светлых линий. Описанные щели носят название вырезок (инцизур) Шмидт-Лантермана (фиг. 9, 1 и фиг. 11) и, по мнению некоторых исследователей, заполнены особенным веществом, отличным от миелина. Шмидт-Лантермановские вырезки заметны на нервных волокнах, взятых от живого животного, напр. лягушки, и рассматриваемых в физиологическом растворе поваренной соли или в других индифферентных жидкостях, а потому вряд ли основательно предположение, высказываемое некоторыми исследователями, что они возникают искусственно, от действия на волокна реактивов или от посмертных изменений в них. Если удалить миелин действием на нервные волокна смеси спирта с эфиром, а затем окрасить их известными красящими веществами, напр. гематоксилином, то нетрудно видеть, что после растворения миелина на месте мякотной оболочки остается густая сеть тонких волокон, сквозь которую ясно просвечивает окрашенный осевой цилиндр. Принимая во внимание, что означенная сеточка не растворима в смеси спирта с эфиром, в щелочах и пр., Эвальд и Кюне причислили ее к так называемой неврокератиновой оболочке (фиг. 8, 2). На самом деле, однако, вещество, из которого состоит эта сеточка, вероятно, представляет собою лишь не растворимые в спирту и эфире примеси, входящие в состав миелина, а потому является образованием искусственным. Снаружи от миелиновой оболочки помещается так называемая Шваннова оболочка, или неврилемма (фиг. 7, b; фиг. 9, 4; фиг. 11, b). Она окружает нервное волокно, нигде не прерываясь, почти от его начала и тянется вплоть до распадания осевого цилиндра на концевые разветвления. В некоторых нервных клетках, к числу которых относятся, напр., спинномозговые и симпатические клетки, означенная оболочка продолжается на поверхность клетки и даже на толстые дендриты и образует вокруг клеточного тела род особенной оболочки или капсулы. Шваннова оболочка кажется в виде тонкой, бесструктурной соединительнотканной пленочки, которая наподобие гильзы окружает нервное волокно и весьма тесно прилегает к миелиновой оболочке. На нервах, обработанных осмиевою кислотой, а затем окрашенных пикрокармином и расщипанных в капле глицерина, в нервных волокнах местами нарушается целость миелиновой оболочки и здесь довольно ясно выступает неврилемма в виде ограничивающих волокна двух тонких линий. Вместе с тем на этих же препаратах заметны еще овальные, окрашенные в розовый цвет ядра, которые тесно прилегают к внутренней поверхности Шванновой оболочки. Означенные ядра называются ядрами неврилеммы (фиг. 8, 3), причем каждое из них окружается небольшим количеством слегка зернистой протоплазмы. Новейшие наблюдения показывают, что от клеток Шванновой ободочки отходят тонкие ветвящиеся отростки, которые соединяются между собою и образуют сеть. По всей вероятности, клетки эти принадлежат к соединительнотканным клеткам и, как показали наблюдения Ранвье, располагаются на известном расстоянии друг от друга; у высших позвоночных, обыкновенно на расстоянии двух перехватов Ранвье, находится лишь одна такая клетка. Некоторые исследователи предполагают, что Шваннова оболочка не покрывает непрерывно все нервное волокно, а окружает лишь каждый отдельный его участок (сегмент) между двумя перехватами, причем на обоих концах участка заворачивается за края мякотной оболочки внутрь и, выстилая внутреннюю поверхность этой оболочки, в то же время непосредственно покрывает собою осевой цилиндр. Указанной части Шванновой оболочки дают название Маутнеровской оболочки или же наружную ее часть называют — наружной, а внутреннюю — внутренней неврилеммой. В мякотных волокнах, входящих в состав периферических нервов, кнаружи от Шванновой оболочки располагается еще особенная соединительнотканная оболочка, так наз. Генлевская оболочка, или эндоневральное влагалище Ретциуса (табл. V, фиг. 1, с). Она состоит из пучков соединительнотканных фибриллей, идущих более или менее параллельно с нервным волокном; к поверхности пучков обыкновенно прилегают отростчатые соединительные клетки, ядра которых легко обнаружить на окрашенных препаратах. Однако на самом деле означенная оболочка не представляет собою отдельного, обособленного для каждого нервного волокна соединительнотканного слоя и, следовательно, совершенно неправильно носит название оболочки. В этом легко убедиться на поперечных разрезах нервных стволиков. На подобных разрезах видно, что нервный стволик окружается эпиневральной оболочкой, в состав которой, кроме пучков соединительнотканных фибриллей и клеток, входят еще сети эластических волокон, отдельные жировые клетки или группы их, а равно кровеносные сосуды, лимфат. щели и канальцы и нервы, так назыв. "nervi nervorum". От этой оболочки отходят различной толщины прослойки, которые проникают в глубину нервных стволиков и разделяют их на известное количество участков — пучков Соединительнотканная прослойка, окружающая каждый такой пучок, носит название периневральной оболочки и в свою очередь посылает от себя тонкие перегородки, разделяющие данный пучок на известное количество более мелких — первичных пучков. Эти перегородки составляют эндоневральную оболочку, причем элементы их — тонкие пучки соединительнотканных фибриллей и клеток — внедряются в промежутки между нервными волокнами, отделяют последние друг от друга и образуют вышеупомянутую Генлевскую оболочку. Самые волокна на поперечных разрезах кажутся в виде кружочков различного диаметра. В центре каждого из них помещается круглое или овальное пятнышко — осевой цилиндр, затем следует более или менее толстое кольцо, то мутное — слоистого строения, то черное — это миелиновая оболочка, а кнаружи от нее находится тонкая линия, обозначающая поперечный разрез Шванновой оболочки. Между нею и миелиновой оболочкой на разрезах некоторых волокон видны в разрезе ядра клеток Шванновой оболочки в форме серпов или полулуний. Промежутки между волокнами заняты фибриллярной соединительной тканью (Генлевская оболочка) с клетками, ядра которых имеют на разрезе круглую или овальную форму. Периневральная оболочка с внутренней своей стороны выстлана плоскими клетками эпителия, границы между которыми на препаратах, обработанных азотнокислым серебром, обозначаются в виде бурых или черных линий. Однако не все мякотные нервные волокна имеют Шванновую и Генлевскую оболочки: волокна белого вещества головного и спинного мозга обыкновенно состоят лишь из осевых цилиндров и окружающей их миелиновой оболочки. Мякотные волокна на своем пути, от начала и вплоть до распадения на концевые веточки, подвергаются делению. Волокна, в которых мякотная и Шванновая оболочка начинаются на известном расстоянии от клетки и которые, следовательно, в начале являются лишь голыми осевыми цилиндрами, обыкновенно отдают от себя известное количество то более, то менее длинных боковых веточек, лишенных оболочек и известных под назв. коллатералей. Начиная с того места, где волокна получают оболочки, они также подвергаются делению на местах перехватов Ранвье. Здесь прежде всего делится Т- или V-образно осевой цилиндр, а затем вскоре каждая из его ветвей получает миелиновую оболочку и таким образом превращается в мякотное волокно. Последнее, пройдя известное расстояние, опять может разделиться на 2 веточки и т. д. Таким путем одно миелиновое волокно нередко дает начало многим подобным же волокнам. Часто на месте одного из перехватов осевой цилиндр делится на 3, 4, 5 и большее количество ветвей, или же от него отходят 1—2 ветви, которые на всем своем протяжении не имеют миелиновой оболочки. Подобного рода деление мякотных волокон постоянно наблюдается на всем их протяжении, пока, наконец, каждое из них не потеряет всех своих оболочек, после чего обыкновенно осевой цилиндр волокна распадается на концевые веточки. Безмякотные, или бледные (Ремаковские), волокна (табл. V, фиг. 2), как показывает само название, отличаются от мякотных волокон, главным образом, отсутствием миелиновой оболочки, отчего они и кажутся не столь резко очерченными, бледными. К ним относятся преимущественно нервные отростки симпатических клеток, почему волокна эти можно бы назвать симпатическими нервными волокнами. Но здесь же необходимо оговориться, что между симпатическими нервными клетками имеются и такие, нервные отростки которых окружаются миелиновой оболочкой. К подобным клеткам принадлежат не только известные клетки симпатической нервной системы человека и млекопитающих, но и многие симпатические клетки низших позвоночных (амфибий, рептилий и др.). В свою очередь, среди клеток спинномозговых узлов млекопитающих также встречаются клетки, нервные отростки которых являются в виде безмякотных волокон. Наконец, в эмбриональной жизни и у молодых животных нервные отростки клеток сначала не имеют мякотной оболочки, которая возникает лишь впоследствии. В состав каждого безмякотного волокна входит осевой цилиндр и окружающая его тонкая, бесструктурная соединительнотканная оболочка — Шванновая оболочка. Осевой цилиндр имеет точно такое же строение, как и в мякотных волокнах, и представляется то в виде толстого волокна, то в форме более или менее тонкой нити, нередко усаженной веретенообразными вздутиями — варикозностями. Что касается Шванновой оболочки, то к внутренней ее поверхности также прилегают звездчатой формы клетки с ядрами (табл. V, фиг. 2, 1), но они расположены на более близком расстоянии друг от друга, чем в мякотных волокнах. Шванновая оболочка, как было сказано выше, часто продолжается на поверхность клетки, напр. в симпатических клетках, и окружает ее в виде особенной оболочки или капсулы, которая продолжается даже на толстые дендриты. Она прекращается лишь вблизи распадения волокон на концевые разветвления. К безмякотным волокнам причисляют неправильно и волокна, входящие в состав обонятельного нерва. Волокна эти имеют вид тонких варикозных ниточек, начинающихся от особенных, так назыв. обонятельных, клеток, и обыкновенно лишены не только миелиновой, но и Шванновой оболочки, т. е. принадлежат к голым осевым цилиндрам. Что касается физиологического значения оболочек, окружающих осевой цилиндр нервного волокна, то некоторые исследователи (Бехтерев, Вестфаль и др.) приписывают миелиновой оболочке роль изолятора, указывая на то, что возбудимость нервных волокон постепенно усиливается по мере образования вокруг них означенной оболочки. Но такое предположение едва ли справедливо, так как безмякотные волокна в течение всей жизни животного остаются лишенными мякоти, а между тем это ничуть не мешает их возбудимости. Кроме того, многие мякотные волокна в начальной своей части и все на месте распадения на концевые разветвления теряют миелиновую оболочку, сохраняя вполне свою возбудимость. Скорее всего, она служит для защиты волокон от различного рода внешних влияний. Разнообразные питательные вещества проникают в мякотные волокна и достигают осевого цилиндра, вероятно, на местах перехватов Ранвье и вырезок Лантерманна.

ТКАНИ ЖИВОТНЫЕ V.
Фиг. 1. Часть поперечного разреза нерва. а — осевой цилиндр; b — мякотная оболочка; прослойки соединительной ткани. Увелич. в 500 раз. Фиг. 2. Безмякотные нервные волокна из стенки желчного пузыря собаки. 1 — ядра Шванновой оболочки. Увелич. в 240 раз. Фиг. 3 и 4. Коротколучистый и длиннолучистый астроциты из мозжечка собаки. Увелич. в 240 (фиг. 3) и в 80 (фиг. 4) раз.
Развитие нервной ткани. Элементы нервной ткани происходят из эктодермы, которая у позвоночных образует сначала впячивание, имеющее вид желобка, а затем постепенно свободные края этого желобка сближаются, наконец сливаются друг с другом и образуют первичную нервную трубку. Довольно толстые стенки этой трубки состоят из многих рядов сначала недифференцированных эктодермальных клеток. Каким образом из этих клеток постепенно возникают различные элементы нервной ткани — это впервые было выяснено наблюдениями Гиса (в 1890—1893 гг.). Он указал, что уже в очень ранней стадии развития зародыша некоторые из клеток мозговой трубки превращаются в круглые, светлые клетки — "зародышевые нервные клетки"; один конец этих клеток постепенно вытягивается по направлению кнаружи и вскоре принимает вид более или менее длинной нити. Таким образом изменившиеся зародышевые нервные клетки получают название "невробластов", а отходящая от каждой из них нить есть первый зачаток нервного отростка — будущего нервного волокна. Вслед за появлением нервного отростка от тела клетки начинают вырастать сначала короткие, похожие на сосочки отпрыски; они постепенно удлиняются, древовидно делятся и в конце концов превращаются в дендриты нервных клеток. Что касается остальных клеток, которым Гис дает название "спонгиобластов", то многие из них впоследствии превращаются в элементы "невроглии", т. е. поддерживающей Т. центральной нервной системы. Наблюдения Гиса подтверждены многими, как, напр., Ленгоссеком, Рециусом, Кахалем и др. Оболочки нервных волокон развиваются лишь позднее, причем относительно способа их развития исследователи до сих пор еще не пришли к определенному заключению. По мнению одних (Ранье, Фюрст), как миелиновая, так и Шванновы оболочки происходят из соединительнотканных клеток, в которых вырабатывается миелин, а остаток протоплазмы с ядром оттесняется к периферии. Осевой цилиндр — нервный отросток, — выйдя из центральной нервной системы, как бы прорастает эти своеобразным способом изменившиеся соединительнотканные клетки, причем каждый промежуток волокна, ограниченный двумя перехватами, отвечает одной такой клетке; оболочка последней составляет Шваннову и Маутнеровскую оболочки нервного волокна, превратившаяся в миелин протоплазма является в виде миелиновой оболочки, а остаток еще не изменившейся протоплазмы вместе с ядром представляется в форме клетки Шванновой оболочки. Однако подобный способ происхождения оболочек нервного волокна не выдерживает критики, так как, с одной стороны, есть мякотные волокна, напр., в центральной нервной системе, не имеющие Шванновой оболочки, с другой — в безмякотных волокнах отсутствует миелиновая оболочка. Ввиду этого пришлось прибегнуть к другим гипотезам образования миелиновой оболочки, к числу которых относится гипотеза, будто означенная оболочка является продуктом, выделяемым осевым цилиндром. При перерезке нервных волокон погибает — дегенерируется — периферический конец их, и восстановление волокон происходит на счет роста, постепенного удлинения осевого цилиндра центрального конца волокна, находящегося в непосредственной связи с нервною клеткою.
Невроглия. К нервной Т. следует еще причислить элементы, служащие для поддержки нервных клеток и волокон в центральной нервной системе, или так назыв. невроглию (глию). Невроглия развивается из тех же элементов, из которых происходят и нервные клетки, т. е. из клеток эктодермы, но не имеет ничего общего с соединительною тканью. К элементам невроглии относятся клетки и волокна. Клетки невроглии встречаются в двух формах: в виде клеток эпендимы и в форме так назыв. астроцитов, или клеток Дейтерса. Первые выстилают как поверхности, так равно и полости центральной нервной системы, напр. центральный спинномозговой канал и пр., и представляются в виде клеток цилиндрической формы, иногда снабженных мерцательными волосками и расположенных то в один, то в несколько рядов. От базального конца каждой клетки обыкновенно отходит один более или менее толстый отросток, который проникает в серое и белое вещество мозга и постепенно распадается на множество ветвящихся нитей. Последние достигают иногда, напр. в спинном мозгу, периферии той или другой части мозга и нередко оканчиваются коническими утолщениями. Означенные волокна называются волокнами эпендимы. Астроциты (табл. V, фиг. 3 и 4) представляют собою маленькие угловатой формы клетки с большим сравнительно ядром, окруженным незначительным количеством протоплазмы. От углов клетки отходит множество тонких, иногда ветвящихся отростков, которые наподобие лучей расходятся в разные стороны и кажутся то короткими, то весьма длинными, вследствие чего и самые клетки получают название коротколучистых (табл. V, фиг. 3) или длиннолучистых (табл. V, фиг. 4) астроцитов. Что касается волокон невроглии, то они имеют различную толщину, пронизывают тела клеток невроглии или идут по их периферии и, по всей вероятности, произошли из клеток. Волокна эти перекрещиваются и переплетаются между собою различным образом и вместе с отростками клеток эпендимы и астроцитов образуют род войлочной массы, в которой заложены нервные элементы, входящие в состав центральной нервной системы.
V. Кровь и лимфа также причисляются, хотя и не совсем верно, к числу простых Т., так как в состав той и другой из них входят жидкость и взвешенные в ней форменные (морфологические) элементы, причем жидкую часть крови и лимфы приравнивают к межклеточному, или основному, веществу поддерживающей Т. Таким образом, кровь и лимфу можно бы назвать "жидкою Т." (подробности — см. Кровь и Лимфа).
А. Догель.
|
Page was updated:Tuesday, 11-Sep-2012 18:16:41 MSK |