| [ начало ] | [ Ц ] |
Центрозома
или центральное тельце — особое тельце, наблюдаемое в большинстве клеток и имеющее, по-видимому, свою определенную функцию в клетке. Относительно природы этого тельца нет вполне установленного взгляда, и некоторые цитологи до сих пор считают Ц. обособившимся участком клеточной протоплазмы вследствие того, что окружающая его протоплазма получает, как увидим, своеобразное расположение. Ц., с этой точки зрения, лишь кажущееся явление, но, однако, по-видимому, за Ц. надо признать некоторую самостоятельность. В редких случаях наблюдали (впервые Брауэр в мужских половых клетках лошадиной аскариды), что Ц. лежит в ядре, почему и приписывать ей происхождение за счет ядра. Так, у одной турбеллярии (Thysanozoon) Ц. появляется в яйце в виде хроматинного волокна и выходит из ядра в протоплазму. Обыкновенно же Ц. является в виде небольшого, лежащего около ядра, сильно преломляющего свет тельца, имеющего свои специфические реакции, вроде энергичного окрашивания железноквасцовым гематоксилином, хотя, по Фишеру (1899), эту же окраску дают и некоторые другие зернистости. Кругом этого тельца наблюдается слой протоплазмы, который носит название центросферы, или сферы притяжения, или архоплазмы. От этой сферы в известные периоды жизни клетки можно отличить отходящие плазматические лучи в виде звезды, или астросферу. Впрочем, терминология этой части клетки не вполне установлена. В окружающей Ц. протоплазме можно отличить два слоя: внутренний, более ясно очерченный на периферии, и наружный, неясно отграниченный от протоплазмы клетки. В применении терминологии к этим частям существует значительное разнообразие. Одни сферой притяжения называют внутренний из упомянутых слоев, а наружный считают непостоянным, чисто протоплазматическим образованием. Другие, наоборот, внутренний слой называют Ц., оставляя за центральными зернистостями название центрального тельца, а за периферическим слоем — сферы притяжения. С точки зрения теории ячеистого строения протоплазмы Ц. рассматривают как одну или несколько ячеек, архоплазму как участок обыкновенной протоплазмы, но лишь с крайне мелкими ячеями, а радиальные волокна звезды как результат слияния вытянутых по радиусам ее ячей. Что касается до Ц. в теле одноклеточных простейших животных, то во многих случаях она обнаружена с полной ясностью. Так, напр., Ишикава наблюдал ее у ночесветки, Шаудин у Paramoeba, загадочного организма, проходящего стадию амебы и потом превращающегося в типичного жгутононосца, а также у солнечников и др. Гортвиг у Асtinosphaerium наблюдал образование Ц. внутри ядра и выхождение ее из ядра в клетку, так что и у простейших она по природе и происхождению принадлежит ядру. Иногда в теле простейших наблюдалось и звездчатое расположение протоплазмы кругом Ц., тогда как в других случаях его не было заметно. В некоторых клетках наблюдаются еще "придаточные ядра". Весьма вероятно, что под этим именем разумеют несколько разнородных образований, природа которых далеко еще не выяснена. Боллес-Ли приходит к заключению, что в одних случаях под этим именем описывались центрозомы с их сферами, которые иногда, по-видимому, могут дегенерировать и служить как бы центром для образования в клетке своеобразных отложений, известных под именем желточных ядер, в яйцах пауков и др., в других случаях под этим именем описывались части хроматина, вытолкнутые ядром в клетку, а в третьих — специально протоплазматические образования и т. п. Перед делением клетки, иногда весьма задолго, Ц. тоже делится, и случается, что еще процесс этого деления не окончен, а Ц. уже разделилась в видах следующего деления клетки. За делением Ц. обыкновенно следует деление архоплазмы и всей астросферы, а потом ядра и клетки. Однако с точностью мы знаем роль Ц. при кариокинетическом делении, а ее роль при прямом делении не выяснена, тогда как при почковании у простейших, по наблюдениям Шаудина, в отпочковывающемся участке Ц. возникает заново. Некоторые приписывают чрезвычайно важную роль Ц. и астросфере при делении клетки. Расположение радиально расходящимися лучами протоплазмы кругом Ц. и ее влияние на деление ядра дали прежде всего идею об электрическом воздействии Ц. на клетку. Галлардо (1896) взял цилиндрический сосуд, наполненный терпентином с кристаллами сернокислого хинина, и на концах его поместил по одному пуговчатому проводнику, стоявшему в сообщении с электростатической машиной. При разрядах последней он получил такое расположение кристаллов хинина около пуговок, какое представляет ахроматинное вещество при непрямом делении, т. е. систему лучей веретена и две системы лучей около сфер притяжения. Идея относительно участия электрических сил при делении клетки была впервые высказана Геннеги и Циглером, но последующие наблюдения над влиянием электрического тока на делящиеся клетки показали, что самое большее, чего мы можем достигнуть, — это удлинения клеток и кариокинетических фигур по направлению тока в местах приложения электродов, и на направление деления ток не влияет. Ряд других авторов, как Ван-Бенеден, Рабль, Бовери, а в позднейшее время Гейденгайн, придают главное значение сокращению лучей, или, по их представлению, фибрилл ахроматинной и плазматической фигуры.
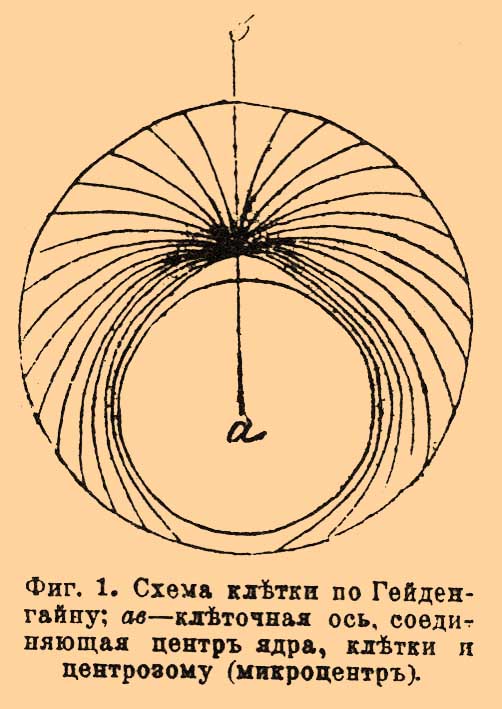
Фиг. 1. Схема клетки по Гейденгайну; ab — клеточная ось, соединяющая центр ядра, клетки и центрозому (микроцентр).
Мы остановимся на воззрениях Гейденгайна, иллюстрируемых придуманным им механизмом. В клетке Гейденгайн отличает ось, соединяющую центр ядра, центр клетки и Ц. (микроцентр) — три точки, лежащие в покоящейся клетке на одной прямой (фиг. 1). Если бы в клетке не было ядра, то Ц. лежала бы в центре клетки, а вследствие присутствия ядра она лежит эксцентрично. К Ц. и к периферии клетки прикрепляются фибриллы, сократимые и не равной длины. При отсутствии ядра они удерживали бы Ц. в центре клетки, но так как ядро сдвигает Ц., то одни из фибрилл в силу их эластичности укорачиваются, другие удлиняются, и наиболее длинные фибриллы, охватывающие кругом ядро, находятся в состоянии наибольшего натяжения. Гейденгайн для воспроизведения этих отношений взял округлый стол. Ц. изображали два связанных кольца. К ним прикрепляли каучуковые шнуры равной длины, причем одна половина шнуров прикреплялась к одному кольцу, другая — к другому, а другим концом все шнуры были укреплены на периферии стола. Положение Ц. было в центре стола. Затем между шнурами вкладывался округлый кусок картона, изображающий ядро.
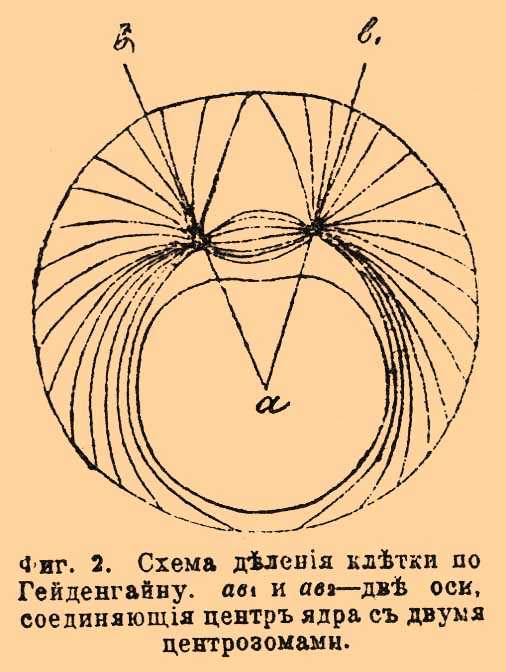
Фиг. 2. Схема деления клетки по Гейденгайну. ab1 и ab2 — две оси, соединяющие центр ядра с двумя центрозомами.
Если два кольца, изображающие Ц., соединить пучком растяжимых нитей, то они, как показывает фиг. 2, под влиянием натяжения каучуковых шнуров разойдутся. Таким образом, это расхождение изображает деление Ц., причем в клетке это расхождение сопровождается, вероятно, разрастанием центрального веретена (см.), соединяющего Ц. Очевидно, что, если бы обе Ц. (в модели кольца) поместились на полюсах ядра (т. е. картона), а ядро в центре веретена (т. е. соединяющих кольца нитей), то тогда клетка достигла бы стадии равновесия и покоя. Но в клетке мы имеем дело не с неподвижной окружностью, как в модели, а с упругой, и натяжение фибрилл не остается без влияния на форму клетки. Если вместо окружности стола, взять окружность из гибкой стали, укрепив внутри ее таким же образом два кольца, соединенных растяжимыми нитями между собой и с периферией кольца, то в силу условий натяжения растяжение колец будет сопровождаться удлинением окружности и даже образованием посредине ее перетяжки, представляющей как бы начало деления самой клетки. Таким образом, деление Ц., ядра и клетки происходит всегда в направлении, перпендикулярном к клеточной оси, и деление, по Гейденгайну, является следствием условий натяжения клеточных фибрилл. Самое натяжение обусловливается положением ядра и натяжением поверхностных слоев клетки вследствие некоторого давления со стороны находящейся в клетке жидкости, или так назыв. тургора (см.). Наконец, третье воззрение, высказанное Бючли и развитое Румблером, ищет причину деления в осмотических явлениях, происходящих в клетке.
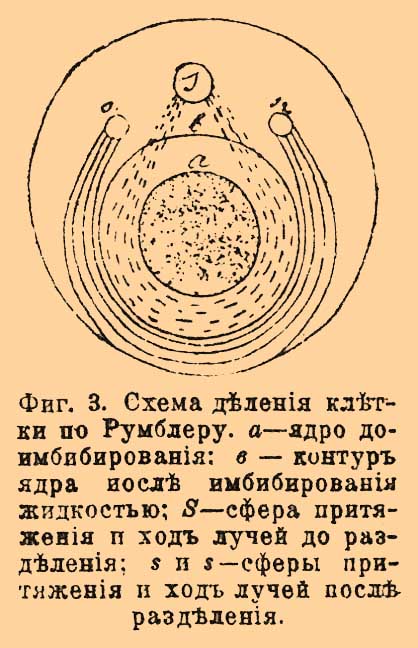
Фиг. 3. Схема деления клетки по Румблеру. a — ядро до имбибирования: b — контур ядра после имбибирования жидкостью; S — сфера притяжения и ход лучей до разделения; s — сферы притяжения и ход лучей после разделения.
Осмотическое перемещение жидкостей, по Румблеру, является главной причиной, вызывающей деление Ц. и ядра. Замечено, что сфера притяжения в начале деления уменьшается, а ядро увеличивается в объеме. Если мы допустим, что лучи протоплазмы, отходящие от сферы (фиг. 3) и образовавшиеся вследствие вышеуказанных осмотических явлений, окружают ядро наподобие кольца, то увеличение объема ядра должно вызвать напряжение этих лучей и разрыв их в месте наиболее эластичном, в области сферы притяжения, что и вызовет разделение сферы и Ц. и расхождение их. Затем наступает увеличение объема сфер притяжения и в силу этого образование около них системы лучей. В силу чисто механических законов, разделившиеся сферы устремляются по направлению лучей наименьшего радиуса, т. е. к полюсам ядра. При таком положении части ядерного веретена, ближайшие к сферам, отдавшие более воды этим последним, будут эластичнее и более вязкими, чем экваториальные части веретена. В силу происходящих осмотических процессов менее эластичные и более богатые водой ячейки притягиваются более эластичными и менее богатыми водой, а следовательно, экваториальные ячейки будут приближаться к полярным, увлекая при этом хромосомы по направлению к двум полюсам. Затем, сферы притяжения снова теряют способность имбибирования водой и уменьшаются, протоплазма принимает равномерную консистенцию и лучистость пропадает. Таким образом, Румблер главным фактором сокращения лучей видит перемещение жидкости или явления осмоса в различных частях клетки. Впрочем, он, как и Гейденгайн, допускает (1897) влияние целого ряда факторов: изменения объема ядра, натяжения лучей, изменения формы клетки перед делением, но в общем его гипотеза является попыткой перенести гипотезу Гейденгайна на почву теории ячеисто-пенистого строения протоплазмы. Возможно деление клетки при отсутствии Ц., а равно при наличности ее, но в отсутствии ядра (см. Физиологическое разделение труда; здесь же разобран вопрос о роли Ц. в отправлениях покоящейся клетки). Что касается до роли Ц. при оплодотворении, то надо различать судьбу Ц. женской, или Ц. яйца, и Ц. мужской, или Ц. живчика. У лошадиной аскариды, судьба мужской Ц., по-видимому, такова: она делится пополам и образует таким образом две Ц., которые ложатся на полюсах образующейся кариокинетической фигуры, окружаются системами радиальных лучей и дают каждая Ц. двух первых дочерних клеток. Бовери, а затем целый ряд исследователей у весьма разнообразных форм наблюдали или допускают исчезновение женской Ц., и большинство их утверждает, что Ц. первой кариокинетической фигуры образуются делением только одной мужской Ц. Фоль описал, что у морских ежей, а равно и некоторых других форм, женская Ц. тоже делится на 2 части, которые сливаются попарно с разделившейся мужской, и потом продукты слияния образуют уже Ц. дочерних клеток. Процесс этот он назвал кадрилью центров, и в этом процессе Ц. каждой клетки дробящегося яйца, как и ядро, имеет двойственное происхождение: мужской и женской. Надо заметить, что наблюдения Фоля нашли подтверждение в наблюдениях Гиньяра над растениями, а также и в наблюдениях над некоторыми животными, напр., Конклина над моллюсками, фан-дер-Штрихта над турбелляриями и др. Наконец, если некоторые не видели настоящей кадрили центров, то все же утверждаюсь, что женская Ц. еще заметна в яйце после выделения второго редукционного пузырька. Таким образом, по-видимому, возможны два случая. Первый случай, когда Ц. оплодотворенного яйца образуются слиянием мужской и женской Ц. Второй случай, когда Ц. оплодотворенного яйца образуются делением одной мужской, причем женская, по некоторым данным, дегенерирует в яйце, а по другим, — всецело выталкивается вместе с редукционным пузырьком. Наконец, наблюдения Уилера (Wheeler) над своеобразным червем Mysostoma заставляют допустить третий случай, а именно, по словам этого автора, живчик Mysostoma не содержит Ц., и Ц. оплодотворенного яйца образуется исключительно за счет женской Ц.. Впрочем, Костанецкий отрицает эти наблюдения и оплодотворение у этого червя относит к типу аскариды. Во всяком случае, первый тип надо считать исходной, более древней формой, а прочие два — его видоизменениями. Если допустить, что Ц. вовсе не имеет никакого отношения к передаче наследственных свойств отца и матери потомству, а является исключительно кинетическим центром (см. Центросфера), то такое разнообразие в происхождении Ц. оплодотворенного яйца не представит ничего парадоксального. При образовании редукционных пузырьков у насекомых Генкинг обнаруживал Ц. и астросферу пузырька действием повышенного атмосферного давления, а Сала — в яйцах аскариды путем их охлаждения, а в других случаях она доступна наблюдению при обыкновенных условиях. Надо думать, что Ц. всегда присутствует в редукционных пузырьках. Ср. Шимкевич, "Биологические основы зоологии" (СПб., 1901); Э. Вильсон, "Роль клетки в развитии и наследственности" (М., 1900, перевод Линдемана), где приведена специальная литература.
В. Шимкевич.
|
Page was updated:Tuesday, 11-Sep-2012 18:16:53 MSK |