| [ начало ] | [ Ч ] |
Черви
или Vermes — установленный Линнеем тип животного царства. В этот тип соединяют самых разнообразных животных, которые по своему строению представляют значительные различия, но более или менее тесно связаны между собой переходными формами, а с другой стороны, обнаруживают родственные отношения к другим типам животного царства. Этот тип подвергался, более всех остальных типов, всяким изменениям, и даже в настоящее время взгляд на то, что мы должны понимать под именем "червей", еще не вполне установился, ввиду обширности, разнообразности и малой характерности типа Ч.; делались даже неоднократно попытки совершенно уничтожить этот тип и разделить его на несколько самостоятельных типов. Характеристика Ч. затрудняется, главным образом, тем 1) что Ч. издавна представляли то "складочное место", в которое относили всех тех животных, которые не находили себе подходящего места среди других типов животного царства и тем 2) что Ч. стоят на рубеже других, более обособленных типов животного царства и, кроме того, еще изобилуют так называемыми переходными формами, т. е. формами, в организации которых отражаются черты строения животных, построенных по разным планам. Это последнее обстоятельство только возвышает интерес изучения этого типа, имеющего громадное значение для эволюционной теории. Ч., как показывают современные сравнительно-анатомические и эмбриологические исследования, произошли от кишечнополостных, с которыми они связаны переходными формами Ctenopl a na и Coeloplana (см. Целоплана), и в свою очередь дали начало всем остальным типам животного царства, за исключением Protozoa, т. е. моллюскам, иглокожим, членистоногим и хордовым или позвоночным. В силу изложенных обстоятельств, при определении характеристики Ч. приходится прибегать к столь общим признакам, что эти признаки могут быть распространены и на другие типы животного. В настоящее время к Ч. относят животных с продолговатым телом, представляющим билатеральную, т. е. двустороннюю, симметрию, с ясно выраженной брюшной и спинной сторонами и передним и задним концом тела. Вторым характерным признаком является присутствие так называемого кожно-мускульного мешка и третьим — наконец, что зародыш образуется из всего желтка, без первоначального появления так называемой первичной бороздки. При такой неопределенности диагностических признаков, границы типа Ч. являются весьма произвольными и разными учеными понимаются различно. Исследования последних лет показали, что тип Ч. может быть разделен на три самостоятельные группы или три подтипа: 1) плоские черви или Platyhelminthes s. Platodes s. Parenchymatosa s. Acoelomata. 2) Настоящие черви или черви в тесном смысле слова — Vermes s. str. и 3) червеобразные или Vermidea. Ввиду того, что первый и третий подтип рассмотрены в соответствующих статьях (см. Плоские черви и Червеобразные) достаточно подробно, мы ограничимся только описанием организации настоящих Ч. или Vermes s. str.
Подтип настоящих Ч. характеризуется удлиненным, двусторонне симметричным телом, однородным по всей своей длине, или же метамерным, т. е. состоящим из однородных метамер или сегментов, лишенных членистых придатков. Далее характерно присутствие кожно-мускульного мешка, кишечника, состоящего из трех отделов и выделительной системы, устроен нос у большинства по типу метанефридий. Подтип Ч. мы подразделяем на следующие 7 классов: 1) Nemertini, 2) круглые черви или Nematodes, 3) колючеголовые или Acanthocephali, 4) Archiannelides, 5) щетинконогие или Chaetopoda, 6) пиявки или Hirudinei и 7) Echiuri d ea. Четыре последних класса соединяют в одну группу кольчатых Ч. или Annulata s. Annelides, которой противопоставляют группу некольчатых Ч. или Anannulata, к которой причисляют Nemertini, Nematodes и Acanthocephali (см. соответствующие статьи, а также Глисты). Настоящие Ч. встречаются в морях и пресных водах, некоторые являются наземными формами (как, например, некоторые Nematodes, Chaetopoda и Hirudinei), a другие ведут паразитический образ жизни, встречаясь внутри растений или животных (Nematodes и Acant h ocephali) или на наружных покровах последних (Hirudinei).
Форма и величина тела весьма разнообразны; у большинства она вытянутая, червеобразная, нитевидная и несколько сплющенная (Nemertini) или цилиндрическая, представляющая круг в поперечном сечении (Nematodes, Acanthocephali), или же цилиндрическая и сплющенная в спинно-брюшном направлении (Annelides). Величина тела также сильно варьирует: встречаются микроскопически малые формы (Nematodes) и достигающие несколько метров длины (Nemertini). У Anannulata тело однородно по всей своей длине, тогда как у Annelides поделено на отдельные метамеры или сегменты, вполне однородные, за исключением первого и последнего сегмента, несущие различные придатки в виде лопастей, щупиков и жаберных нитей (у Chaetopoda Polyc h aeta Sedentaria). У Chaetopoda на каждом членике с обеих сторон встречаются по две пары щетинок, сидящих в особых углублениях или кармашках и приводимых в движение особыми мышцами, или же с каждой стороны по выступу, несущему по целому пучку щетинок (табл., фиг. 6) и щупику с жаберными придатками — так называемые параподии, являющиеся примитивными органами передвижения. У других Ч. встречается на переднем конце тела придаток, так называемый хоботок (табл., фиг.1 p), снабженный стилетом у некоторых Nemertini или несколькими венчиками крючьев у Acanthocephali. Этот хоботок может быть втягиваем внутрь тела в так называемое влагалище (табл., фиг. 1 ps) или же выпячиваем наружу и служит им или как орган защиты, или как орган прикрепления. У пиявок, наконец, на переднем и заднем конце тела встречается по присоске (табл., фиг. 3), при помощи которых они присасываются к животным, на которых паразитируют.
Покровы Ч. состоят из однослойного эпидермиса (табл., фиг. 4е) эктодермального происхождения, в котором различают обыкновенные цилиндрические или опорные клетки, затем железистые, пигментные и веретенообразные чувствительные клетки. У Nemertini эпидермис покрыт мерцательными ресничками, тогда как у прочих Ч. одет более или менее толстой, стекловидной кутикулой (табл., фиг. 4 ct), представляющей продукт выделения под ней лежащего эпидермиса. Эта кутикула сбрасывается при линьке и заменяется новой. Особенной толщины кутикула достигает у паразитических Nematodes и Acanthocephali. У взрослых Nematodes эпидермис превращается в мелкозернистый, так называемый гиподермический слой, в котором нельзя различить стенок клеток, а также и ядер, и который на всем своем протяжении образует четыре продольных выступа, вдающихся в виде четырех продольных валиков в полость тела. Под эпидермисом у Nemertini залегает еще тонкий соединительно-тканный слой — так называемая основная или базальная перепонка. Под ней, а у всех прочих Ч. непосредственно под эпидермисом, находятся мышцы, которые, будучи тесно связаны с эпидермисом, представляют характерное для Ч. образование — так называемый кожно-мускульный мешок, сокращениями которого обуславливаются те особые своеобразные движения, которые обыкновенно называются червеобразными. Этот кожно-мускульный мешок состоит обыкновенно из нескольких различной толщины слоев мышц. Так, у Nemertini мы различаем последовательные слои кольцевых, продольных и кольцевых мышц; у Acanthocephali, Chaetopoda, Hirudinei и Echiuridea — слои кольцевых (табл., фиг. 4 и 6 mc) и продольных (ml), иногда своеобразно сгруппированных мышц; а у Archiannelides и Nematodes лишь один слой продольных мышц, которые у последнего класса отличаются весьма своеобразным строением и расположением. У Hirudinei встречаются еще спинно-брюшные мышцы (табл., фиг. 4 mdv), напоминающие таковые же мышцы плоских Ч. За кожно-мускульным мешком следует тонкий эпителий мезодермального происхождения, выстилающий деломическую, т. е. вторичную полость тела.
Полость тела имеется у всех Ч. (табл., фиг. 5 cl), но достигает различного развития. У Nemertini, как показали новейшие исследования, образуется настоящая целомическая полость, но во время последующего развития она заполняется мезодермальными клетками, так что от полости тела остаются небольшие пространства, окружающие хоботок и представляющие его влагалище (табл., фиг. 1 ps), a y некоторых форм еще и кишечник. То же самое наблюдается и у Hirudinei, у которых полость тела во время развития заполняется паренхиматозными клетками (табл., фиг. 4 pr), так что от нее остаются небольшие участки — так называемые синусы (табл., фиг. 4 sl, sd, sv) в которых находится кровь и залегают некоторые органы как напр. нервная и (табл., фиг. 4 s) кровеносная система. У Nematodes Acanthocephali и Echiuridea имеется одна большая полость тела, в которой помещаются различные органы, прикрепленные к ее стенке при помощи особых перегородок или мезентерий мезодермального происхождения, У Archiannelides и Chaetopoda (табл., фиг. 2, 3 и 5), наконец, полость тела поделена поперечными перегородками или септами на отдельные камеры, соответствующими наружным сегментам, а каждая камера продольным брюшным и спинным мезентерием, служащим для прикрепления кишечника на две камеры, соответствующие так называемым первичным сегментам.
Органы пищеварения представлены кишечником, открывающимся наружу ротовым и анальным отверстием и состоящим из передней, средней и задней кишки. Ротовое отверстие помещается на переднем конце тела или в передней части тела (табл., фиг. 1 о), на ее брюшной стороне (Nemertini), a анальное отверстие (табл., фиг. 1 а) на заднем конце тела или же на спинной стороне (Hirudinei). В передней кишке эктодермального происхождения различают обыкновенно пищевод и мускульную глотку, а у некоторых Ч. еще ротовую полость и жевательный желудок. В переднюю кишку открываются различные железы, как, например, так называемые слюнные, известковые и ядовитые железы, а также у некоторых Ч. помещаются особые хитиновые приспособления, как шипы (Nematodes), крючья, челюсти (Chaetopoda и Hirudinei), служащие для ранения, захвата и измельчения добычи. Средняя кишка, энтодермального (табл., фиг. 1, 4, 6 i) происхождения, отличается более значительной длиной и служит для всасывания заглотанной пищи; у Nemertini она образует по всей длине парные выступы, чередующиеся с сужениями кишечника (табл., фиг. 1 i) и представляющие зачаток метамерии, а у Hirudinei имеются парные мешкообразные слепые выросты; в среднюю кишку Chaetopoda открываются одноклеточные железы, выделяющие вещество, играющее известную роль при пищеварении. Задняя кишка эктодермального происхождения обыкновенно коротка; в нее открываются у Hirudinei одна пара так называемых анальных желез, а у Echiuridea два мешка, усаженных мерцательными воронками и являющиеся органами выделения и дыхания. У самцов Nematodes в заднюю кишку открывается еще выводной проток мужских половых органов, почему задний конец задней кишки носит название клоаки. У Acanthocephali кишечник, а равно и ротовое и анальное отверстия отсутствуют совершенно; они утратили его вследствие эндопаразитического образа жизни, питаясь осмотическим путем всей, поверхностью тела.
Нервная система (табл., фиг. 1, 2, 3 n) представляет различные ступени совершенствования у различных классов Ч. Мы различаем в ней: 1) надглоточные ганглии, 2) окологлоточное нервное кольцо, 3) продольный нервный ствол или стволы, тянущиеся от окологлоточного кольца вплоть до заднего конца тела и 4) нервные ветви. У Nemertini имеются два больших ганглия (табл., фиг. 1 ng), соединенных окологлоточной комиссурой, от которой отходят к переднему концу нервы к органам чувств (c, s), а к заднему— 3 нервных ствола, два боковых (табл., фиг. 1 n) и один брюшной, соединенные между собой поперечными комиссурами. У Nematodes от окологлоточного нервного кольца отходят два продольных, брюшной и спинной, нервных ствола, не разветвляющихся на веточки и соединенных между собой поперечными комиссурами. К этим продольным стволам направляются отростки мышечных клеток, представляющие очень своеобразное и единственное в животном мире явление. У Acanthocephali нервная система редуцирована и состоит из нервного узла, залегающего на дне хоботного влагалища и от которого вперед и назад отходят по два нервных ствола. Нервная система Annelides очень интересна в сравнительно-анатомическом отношении. Она состоит из двух надглоточных и подглоточных ганглиев, соединенных кольцевой комиссурой, от которой отходят два нервных ствола, тянущихся на брюшной стороне вдоль всего тела вплоть до заднего конца. Эти стволы у Archiannellides однородны, тогда как у Chaetopoda и Hirudinei снабжены в каждом сегменте ганглиозными вздутиями, соединенными между собой комиссурами и образующие в совокупности брюшную нервную лестницу. При укорачивании комиссур, у высших представителей этих классов, происходит сближение нервных стволов и, наконец, полное слияние в один ствол — так называемую брюшную нервную цепочку (табл., фиг. 2, 3 n). Не менее интересно и положение нервной системы у Annelides. Она образуется в эмбриональной жизни Ч. из эктодермы и у Archiannelides, а также у некоторых представителей Chaetopoda сохраняет это эпителиальное положение на всю жизнь, залегая в эпидермисе. У других высших Chaetopoda и Hirudinei — она перемещается впоследствии в полость тела, где и залегает у взрослых животных (табл., фиг. 4, 6 n). У Echiuridea нервный ствол лишен ганглиозных утолщений — но это явление регрессивное, так как в личиночном состоянии у них закладываются два нервных ствола, снабженные в каждом сегменте ганглиозными утолщениями, которые, сливаясь в один ствол при дальнейшем развитии, а именно при исчезновении перегородок, теряют ганглиозные вздутия и оказываются состоящими по всей своей длине из ганглиозных клеток и нервных волокон.
ЧЕРВИ.
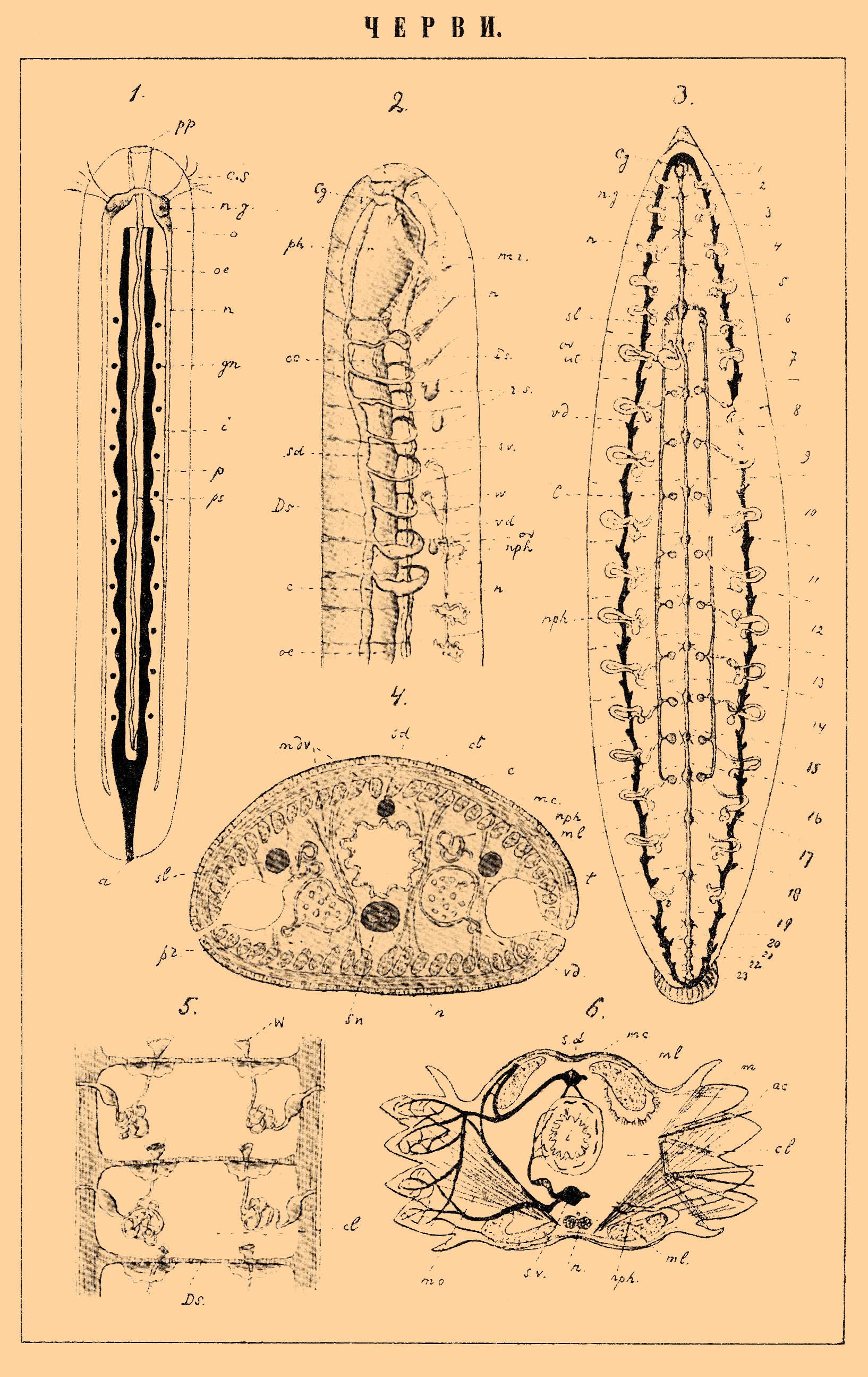
К табл. ЧЕРВИ.
1. Схематическое изображение немертины.
2. Pontodrilus (Chaetopoda Oligochaeta), передний конец тела, вскрытый со спинной стороны.
3. Медицинская пиявка (Hirudo medicinalis); схематизированное изображение нервной, кровеносной, выделительной и половой систем. Цифрами 1-23 обозначены сегменты, границы между которыми показаны пунктирными линиями.
4. То же. Поперечный разрез через тело; схематизировано.
5. Схема расположения выделительных органов (метанефридий) у кольчатого червя (Annelides).
6. Схематизированный поперечный разрез через тело Nereis (Chaetopoda Polychaeta). На левой стороне изображены преимущественно сосуды кровеносной системы, на правой же — щетинки, мышцы и нервы.
Значение букв: a - порошица; ac - главные (опорные) щетинки; c - сердце; Cg — надглоточный ганглий; cl — вторичная полость тела (цёлом); сs — мерцательная ямка; ct — кутикула; Ds — поперечные перегородки (септы); e — эпидермис; gn — половые органы; i — кишечник; m — мышцы; mc — кольцевые мышцы; mdv — спинно-брюшные мышцы; ml — продольные мышцы; mo — косые мышцы, mr — мышца-ретрактор; n — нервный ствол; ng — нервный ганглий; nph — выделительные органы; o — рот; oe — пищевод; ov — яичник; p — хоботок; ph — глотка; pp — отверстие хоботка; pr — паренхима; ps — влагалище хоботка; rs — семяприемник (receptaculum seminis); sd — спинной кровеносный сосуд; sl — боковой кровеносный сосуд; sv — брюшной кровеносный сосуд; t — семенники; ut — матка; vd — vas deferens; W — воронка выделительной системы; w — воронка выводных протоков мужских половых органов.
Органы чувств весьма разнообразны у разных классов Ч. Органы осязания состоят из веретенообразных клеток, помещающихся в виде точек (накожные органы) на разных частях тела, по преимуществу на передней части (у Nemertini, Nematodes), на голове (у Hirudinei) и на щупальцах (у Archiannelides и Chaetopoda). Органы вкуса имеют вид бокаловидных органов и встречаются в глотке и пищеводе Chaetopoda. Органы обоняния устроены в виде мерцательных ямок (табл., фиг. 1 cs), помещающихся у Nemertini и Archiannelides в переднем конце тела. Органы слуха мало распространены среди Ч. и встречаются у некоторых Nemertini и Chaetopoda Polychaeta; они имеют вид маленьких пузырьков, наполненных жидкостью и содержащих один или несколько отолитов из углекислой извести; пузырьки эти поддерживаются клетками, снабженными волосками и сообщающимися с нервной системой. Глаза более распространены у Ч. и встречаются у Nemertini, свободно живущих Nematodes, Chaetopoda и Hirudinei; они располагаются в виде одной или нескольких пар в переднем конце тела и устроены или по типу простых глазных пятен, или по типу камеры-обскуры (у высших Chaetopoda Polychaeta Errantia). Глаза Nemertini и Hirudinei отличаются весьма своеобразным строением. Паразитические Nematodes, равно как Acanthocephali, лишены органов чувств, что, несомненно, находится в связи с их энтопаразитическим образом жизни.
Кровеносная система встречается у всех Ч., за исключением Nematodes и Acanthocephali. У Nemertini она состоит из трех продольных стволов, двух боковых и одного спинного, идущих вдоль всего тела от переднего вплоть до заднего конца; на переднем и заднем конце тела два боковых сосуда соединяются между собой и со спинным сосудом, а у некоторых немертин существует еще несколько поперечных каналов или комиссур. Стенки каналов мускулисты и сократимы, так что кровь движется в них в определенном направлении. У Ann e lides кровеносная система достигает высшей степени развитии; у Archiannelides и Chaetopoda различают один спинной и один брюшной сосуд (табл., фиг. 2, 4, sd, sv), соединяющиеся в каждом сегменте между собой при помощи дугообразных боковых сосудов или комиссур; некоторые из этих сосудов развиты сильнее и пульсируют, а поэтому принимаются некоторыми учеными за сердца (табл., фиг. 2 с). От главных сосудов отходят мелкие кровеносные сосуды, направляющиеся в различные органы. У Hirudinei кровеносные (табл., фиг. 3, 4, sd, sv, sb) сосуды (обыкновенно 2 или 4 продольных) заключены в лакуны, так называемые синусы, представляющие остаток вторичной полости тела, и также сообщаются между собой при помощи комиссур. Наконец, у Echiuridea лишь брюшной кровеносный сосуд тянется во всю длину тела, тогда как сообщающийся с ним спинной сосуд (который принимают также за сердце) идет лишь вдоль передней кишки. Кровь Ч. большей частью бесцветная, а у некоторых Annelides бывает диффузно окрашена в желтоватый, зеленоватый или красный цвет; встречаются форменные элементы крови, которые большей частью бесцветны, хотя у Nemertini было доказано присутствие гемоглобина.
Органы дыхания встречаются у немногих Ч.; большинство Ч. дышат всей поверхностью тела. У Archiannelides окисление крови происходит в щупальцах, а у большинства Chaetopoda Polychaeta имеются особо дифференцированные органы дыхания или жабры; последние у Sedentaria помещаются на переднем конце тела в виде пучка тонких нитей, а у Errantia с боков тела на параподиях (табл., фиг. 6), в виде тонкостенных перистых или кустистых отростков, в которые продолжается полость тела, наполненная кровью. Такие жабры встречаются и у Hirudinei, а именно лишь у одного рода Branchelion. У Echiuridea органы дыхания представлены двумя мешками, выростами задней кишки, которые через заднепроходное отверстие наполняются от времени до времени водой.
Органы выделения Ч. весьма разнообразны и у большинства устроены по типу метанефридий. У Nemertini они имеют еще примитивный характер и отчасти напоминают выделительную систему плоских червей (см.); они состоят из двух продольных каналов, расположенных по бокам в передней части тела или по всей его длине и выстланных мерцательным эпителием; эти каналы разветвляются, причем ветви оканчиваются в паренхиме небольшими расширениями, покрытыми мерцательными ресничками; наружу каналы открываются одним, двумя или несколькими отверстиями на заднем конце или с боков тела. Для некоторых немертин (Palaeonemertini) было доказано непосредственное сообщение выделительной системы с кровеносной, к которой она тесно прилегает, с помощью 2-3 отверстий. У Nematodes выделительные органы еще малоисследованны и состоят из двух продольных каналов, залегающих в боковых линиях (выступах гиподермиса) и сообщающихся между собой в переднем конце тела кольцевым каналом, открывающимся на брюшной стороне небольшим отверстием наружу. В связи с этими каналами стоят, по-видимому, большие клетки, обладающие фагоцитарной функцией и которые помещаются в полости тела, прилегая к боковым линиям. Выделительные органы Acanthocephali неизвестны. У Annelides, т. е. у Archiannelides, Chaetopoda и Hirudinei встречаются так называемые сегментальные органы (табл., фиг. 2-6 nph) или метанефридии; они располагаются попарно в каждом сегменте (за исключением немногих передних) так, что своими широкими, покрытыми ресничками воронками (табл., фиг. 5 w) открываются в полость тела (el), затем переходят в узкий канал, который, прободая перегородку или септу (Ds), образует несколько петлеобразных заворотов (клубов) в следующем сегменте и открывается сбоку тела наружу, образуя у некоторых Ч. небольшое расширение. Такие метанефридии развиваются из протонефридий, встречающихся у этих классов Ч. лишь во время личиночной жизни и остающихся таковыми на всю жизнь у плоских червей (см.). Метанефридии служат не только для выведения наружу последних продуктов обмена веществ, но у многих Chaetopoda и для выведения половых элементов. Наконец, у Echiuridea выделительные органы представлены двоякими образованиями: 1) 1-3 парами мешков, лежащими в переднем конце тела и открывающимися большой воронкой в полость тела и небольшим отверстием наружу, и 2) мерцательными воронками, сидящими на стебельках на больших мешках, открывающихся в заднюю кишку и исполняющих помимо экскреторной функции еще и дыхательную.
Фагоцитарные органы были найдены в последнее время у Chaetopoda; они располагаются попарно в каждом сегменте и прилегают к спинной стороне средней кишки, будучи наполнены амебоидными клетками, заглатывающими всякие посторонние тела, находящиеся в полости тела Ч.
Половые органы Ч. устроены чрезвычайно разнообразно. Проще всего они устроены у высших Ч., т. е. у Archiannelides, Chaetapoda Polychaeta и Echiuridea; y первых двух как мужские, так и женские половые элементы образуются из клеток эндотелия мезодермального происхождения, выстилающего полость тела и обыкновенно на поперечных или трансверсальных перегородках или септах, причем они выводятся наружу при помощи выделительных органов. У Echiuridea половые продукты развиваются также из клеток эндотелия, выстилающего полость тела, но в определенном месте — у одних форм в переднем, у других в заднем конце тела. У Chaetopoda Oligochaeta половые продукты, хотя развиваются из тех же клеток, но в определенных сегментах и на поперечных септах, образуя семенники и яичники, при этом септы у самцов разрастаются и образуют так называемые семенные мешки или резервуары, из которых сперматозоиды заглатываются мерцающими воронками (табл., фиг. 2 w, vd) и выводятся наружу выводными каналами или vas deferens. Яйца, созревая, падают в полость тела (ov) и выводятся наружу трубчатыми яйцеводами, прикрепленными к септе следующего сегмента. Кроме того, имеются еще 2 пары семяприемников (табл., фиг. 2 r, s), служащие для восприятия сперматозоидов при совокуплении от другого неделимого. Половые органы Nemertini устроены так же просто в виде небольших мешочков, расположенных в паренхиме тела по обе стороны средней кишки между ее боковыми выступами (табл., фиг. 1, gn); у других же они разбросаны без всякого порядка по всему телу. Половые продукты образуются в этих мешочках и выходят наружу через выводные протоки, открывающиеся наружу сбоку по обе стороны тела. У Nematodes половые органы имеют вид длинной трубки, парной у самки и непарной у самца, залегающей в полости тела и опутывающей кишечник бесчисленными петлеобразными заворотами. В этих трубках различают следующие последовательно друг за другом лежащие отделы: у самки — яичники (ovarii), яйцеводы (oviducti), семяприемники (receptacula seminis), матка (uteri) и влагалище (vagina); у самца — семенник (testis), семяпровод (vas deferens) и семяизвергательный канал (ductus ejaculatorius) с копуляционным органом (spiculum), имеющим вид щетинки. Женские половые органы открываются наружу самостоятельным отверстием в передней трети тела, а мужские — в задний отдел задней кишки. Половые органы Acanthocephali и Hirudinei в общем устроены по тому же типу. У первого класса мужские половые органы состоят из парных семенников, от которых отходят два выводных протока, соединяющихся в непарный семяизвергательный канал, принимающий несколько предстательных железок (prostata) и переходящий в копуляционную сумку с хитиновым копуляционным органом (penis). Женские половые органы Acanthocephali состоят из яичника, в котором развиваются яйца и, после созревания, выпадают с разрывом стенок яичника в полость тела, где они заглатываются так называемым маточным колоколом и при помощи яйцеводов попадают в матку, а из нее через влагалище наружу. У Hirudinei имеется несколько пар семенников (табл., фиг. 3, 4 t), от которых отходят семяпроводы, впадающие с каждой стороны в один продольный проток (vd); эти протоки соединяются в непарный проток — семяизвергательный канал, который принимает предстательную железу и на брюшной стороне открывается наружу, будучи снабжен еще особым хитиновым копуляционным органом; женские половые органы (табл., фиг. 3 ov) состоят из одной пары яичников, двух яйцеводов, соединяющихся в непарную матку (ut), принимающую так называемую белковую железу и переходящую во влагалище, которое также на брюшной стороне тела, но позади мужского полового отверстия открывается наружу. Громадное большинство Ч. является раздельнополыми; обоеполыми или гермафродитами являются немногие Nemertini, Archiannelides, Chaetopoda Oligochaeta и немногие Polychaeta и Hirudinei. Среди Echiuridea и Chaetopoda Polychaeta встречается даже диморфизм, а у самцов последних еще брачный наряд. Помимо полового размножения, у Chaetopoda встречается еще бесполое — в виде деления и почкования, а также возможна в широкой степени и регенерация.
Эмбриональное развитие Ч. весьма разнообразно и интересно с филогенетической точки зрения. Только у немногих (Nematodes, Chaetopoda Oligochaeta и Hirudinei) имеется прямое развитие, у громадного большинства встречается же метаморфоз и очень интересные личинки — пилидий у Nemertini и трохофора у кольчатых червей (см. соответствующие слова). У паразитических Nematodes и Acanthocepliali мы встречаем перемену хозяина, а у первых еще гетерогонию. Что касается родственных отношений Ч., то Nemertini примыкают к плоским червям, кольчатые черви являются родоначальниками червеобразных (см.), а с ними вместе и других типов животного царства, за исключением Protozoa и Coelenterata. Nematodes и Acanthocephali занимают совершенно обособленное положение; их родственное отношение, особенно последнего класса к другим Ч., установить довольно трудно вследствие регресса, который они претерпели благодаря своему паразитическому образу жизни.
В. Шевяков.
|
Page was updated:Tuesday, 11-Sep-2012 18:16:56 MSK |